東京大学大学院 農学生命科学研究科 プレスリリース
植物細胞のエンドサイトーシス小胞形成に関与する分子装置の特性を解明
- 発表者
- 藤本 優 (東京大学大学院農学生命科学研究科 生産・環境生物学専攻 特任研究員(当時);
現東京大学大学院理学系研究科 生物科学専攻 特任研究員)有村 慎一 (東京大学大学院農学生命科学研究科 生産・環境生物学専攻 助教)
上田 貴志 (東京大学大学院理学系研究科 生物科学専攻 准教授)
高梨 秀樹 (東京大学大学院農学生命科学研究科 生産・環境生物学専攻 博士課程3年)
林 嘉禾 (東京大学大学院農学部 応用生物学専修4年)
中野 明彦 (東京大学大学院理学系研究科 生物科学専攻 教授,理化学研究所 基幹研究所 主任研究員)
堤 伸浩 (東京大学大学院農学生命科学研究科 生産・環境生物学専攻 教授)
発表概要
植物(シロイヌナズナ(注1))のエンドサイトーシス(注2)小胞形成に関与する分子装置には,少なくとも2種類の生体膜切断分子が含まれていることが分かりました.
発表内容
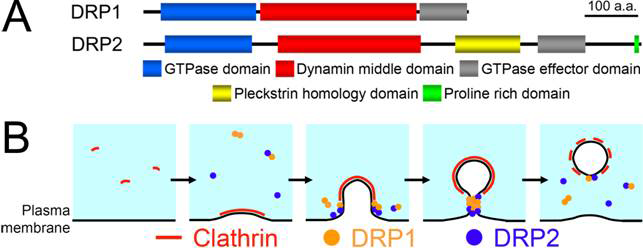
図A シロイヌナズナダイナミン様タンパク質DRP1とDRP2の分子構造
膜脂質への結合に重要なPleckstrin homology domainや小胞形成部位への局在化に重要なProline rich domainはDRP2にのみ存在する
図B シロイヌナズナダイナミン様タンパク質DRP1とDRP2の分子構造
本研究の成果から予想される植物エンドサイトーシス小胞形成時の細胞膜切断機構の一例
エンドサイトーシスとは,真核細胞が外部からの物質の取り込みを行う現象であり,また細胞膜上の脂質やタンパク質の量・配置の調節を行う上で必須の現象です.近年,植物の養分吸収や糖蓄積,病原菌への抵抗といった形質へのエンドサイトーシスの関与が示され,その分子レベルにおける制御機構の解明は重要な課題となっています.そこで我々は,エンドサイトーシスの初期段階である細胞膜からの小胞形成とその切り取りについて,植物における分子装置とその仕組みを解明するために,細胞表面領域の蛍光分子を特異的に可視化することが可能な全反射照明蛍光顕微鏡(注3)とライブセルイメージング(注4)技術を用いてこの課題に取り組みました.
真核細胞内の膜系ダイナミクスを制御する分子の一つに,ダイナミン様タンパク質という分子が存在します.このダイナミン様タンパク質は,自己重合してリングもしくは螺旋状のポリマーを形成して生体膜に巻きつき,その切断を実行すると考えられています.モデル植物であるシロイヌナズナのゲノム中には,DRP1~DRP6という6種のダイナミン様タンパク質が存在することが報告されていました.
植物細胞のエンドサイトーシスにおいては,細胞膜から出芽する小胞の大半がクラスリン(注5)というタンパク質によって覆われていることが分かっています.そこで我々は,まずシロイヌナズナにおいて,エンドサイトーシス小胞形成に関与するダイナミン様タンパク質の有無を調べるため,細胞膜上でこのクラスリンで覆われた場所に出現するような分子種があるかどうかを探索しました.
その結果,DRP1とDRP2というダイナミン様タンパク質が細胞膜上でクラスリン被覆小胞が形成された場所に蓄積することが分かりました.また,これら2種のダイナミン様タンパク質の分子構造は大きく異なるにも関わらず(図A参照),それらの細胞膜上での局在パターンと挙動はほぼ一致し,さらに結合する可能性があることも明らかにしました.このことは,植物のエンドサイトーシス小胞の形成には,構造的に異なる2種類のダイナミン様タンパク質が協調的に機能している可能性を示唆しています(図B参照).
本研究で明らかにした構造の異なる複数種の生体膜切断分子が同一の小胞形成に関与するという概念は新しいものであり,今後さらに,細胞膜の構造変換に果たすDRP1とDRP2分子の機能を解析することで,新たな生体膜切断メカニズムの存在を提示することができるものと期待されます.また,これら分子の機能を人為的に操作することができれば,将来的に養分吸収や糖蓄積といった作物の特性の向上につながることが期待されます.
なお本研究は,文部科学省科学研究費補助金による支援を受けました.
発表雑誌
Arabidopsis dynamin-related proteins DRP2B and DRP1A participate together in clathrin-coated vesicle formation during endocytosis.
<著者名>
Masaru Fujimoto, Shin-ichi Arimura, Takashi Ueda, Hideki Takanashi, Yoshikazu Hayashi, Akihiko Nakano and Nobuhiro Tsutsumi
<公表雑誌>
米国科学アカデミー紀要電子版(Proceedings of the National Academy of Sciences of the United States of America)3月16日号
問い合わせ先
植物分子遺伝学研究室
教授 堤 伸浩
Tel: 03-5841-5073
FAX: 03-5841-5183
E-mail: atsutsu@mail.ecc.u-tokyo.ac.jp
URL: http://park.itc.u-tokyo.ac.jp/pmg/
用語解説
アブラナ科シロイヌナズナ属の一年草.生活環が短い,形質転換が容易であることなどからモデル植物として用いられており,植物としては初めて全ゲノムが解読された.
注2: エンドサイトーシス細胞が細胞外や細胞膜上の物質を取り込む現象の一つ.このエンドサイトーシスの過程では,まず細胞膜が内側に向かって陥入し,袋状の構造が形成され,最終的に袋の根元の膜がくびり切られることによって小胞が形成される.この小胞はエンドサイトーシス小胞と呼ばれ,その後標的となる細胞内小器官へ輸送される.従来,植物細胞では,細胞壁が存在するためにエンドサイトーシスによる細胞外物質の取り込みはないと考えられてきたが,最近の研究では,植物もエンドサイトーシスをおこなっていることがわかっている.
注3: 全反射照明蛍光顕微鏡試料表面付近の蛍光のみを特異的に観察することのできる特殊な顕微鏡.入射光をカバーグラスで全反射させた際に生じるエバネッセント光を励起光として用いる.これにより,細胞内部からのノイズのない細胞膜付近の鮮明な画像を得ることができる.
注4: ライブセルイメージング細胞や組織などの生体材料を固定せず生きたまま観察すること.
注5: クラスリンエンドサイトーシス小胞の形成に重要な分子構造の一つ.このクラスリンは重鎖,軽鎖と呼ばれる分子からなる三角巴構造であり,エンドサイトーシスの際にはそれらが重合して小胞を取り囲むような格子上の籠が組み立てられる.