疑似リン酸化型OsWRKY53転写因子によるイネの耐病性向上に成功
- 発表者
-
中条 哲也(東京大学生物生産工学研究センター プロジェクト研究員;当時)
宮本 皓司(東京大学生物生産工学研究センター プロジェクト研究員;当時)
小川 哲史(東京大学生物生産工学研究センター 博士課程2年)
増田 優花(東京大学生物生産工学研究センター 修士課程2年;当時)
清水 崇史(東京大学生物生産工学研究センター プロジェクト研究員;当時)
岸(加星) 光子(農業生物資源研究所 研究員)
高橋 章(農業生物資源研究所 研究員)
西澤 洋子(農業生物資源研究所 研究員)
南 栄一(農業生物資源研究所 研究員)
野尻 秀昭(東京大学生物生産工学研究センター 教授)
山根 久和(帝京大学理工学部バイオサイエンス学科 教授、東京大学生物生産工学研究センター 教授;当時)
岡田 憲典(東京大学生物生産工学研究センター 准教授)
発表のポイント
◆イネの病害応答性転写因子OsWRKY53の疑似リン酸体の過剰発現が、イネの耐病性を増幅することを発見しました。
◆これまでにOsWRKY53の過剰発現でイネに耐病性を付与すること示されていましたが、MAPキナーゼによるOsWRKY53のリン酸化が耐病性の発揮に重要であることが示唆されました。
◆本発見は、イネの病害抵抗性発現の分子メカニズムの解明につながるもので、農薬使用の少ない環境保全型の農業技術開発への応用が期待されます。
発表概要
東京大学生物生産工学研究センター山根久和教授(当時)と岡田憲典准教授(当時助教)らの研究グループは、いもち病(注1)の原因菌であるイネいもち菌感染時に発現する転写因子OsWRKY53のリン酸化が、イネの耐病性発現に重要であることを明らかにしました。同グループはOsWRKY53がイネの耐病性を増加させることを以前の研究で報告していましたが、いもち菌感染時にイネがOsWRKY53を介してどのように抵抗性を発揮しているのかは不明でした。また、イネの病害抵抗性発現においては、リン酸化シグナル伝達の重要性が広く認められていましたが、OsWRKY53の機能発現とリン酸化の関係についても未解明でした。
今回、イネのMAPキナーゼカスケード(注2)のひとつであるOsMKK4-OsMPK3/OsMPK6によるリン酸化で、OsWRKY53の転写活性化能が上昇することを明らかにし、このリン酸化により病害抵抗性に関与する遺伝子群の発現が促されることを示唆しました。また、OsWRKY53のN末端側に存在するリン酸化部位のアミノ酸のセリンをアスパラギン酸に置換した疑似リン酸化OsWRKY53を用いることで、MAPキナーゼによる活性化と同様にOsWRKY53に転写活性化能を付与できることを見出し、その過剰発現によりイネの耐病性を増強することに成功しました。本成果は、転写因子を利用した新たな病害抵抗性イネの育種技術に発展することが期待されます。
発表内容
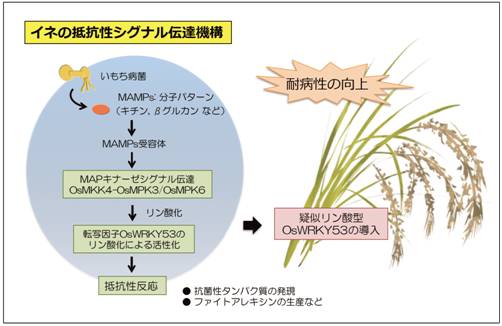
図1 イネにおけるいもち菌感染から抵抗性反応までのシグナル伝達の模式図(拡大画像↗)
転写因子OsWRKY53はOsMKK4-OsMPK3/PsMPK6によりリン酸化を受ける。転写活性化能を保持した疑似リン酸化型OsWRKY53をイネで過剰発現させると、抗菌性タンパク質の発現などを伴い、耐病性の向上が起こる。
植物は病原菌からの攻撃に対し2つの自然免疫系を駆動して対抗しています。ひとつは病原菌特有の分子パターン(注3:MAMPs /PAMPsと総称される)であるキチンオリゴ糖やフラジェリン、リポ多糖などのエリシター(注4)により誘導される基礎的抵抗性であり、もうひとつは病原体由来の非病原性遺伝子産物(AVRタンパク質)と植物由来の抵抗性遺伝子産物(Rタンパク質)との相互作用により誘導される特異的抵抗性です。特異的抵抗性が、ある種の病原菌と植物との間でのみで起こる強い免疫反応であるのに対し、基礎的抵抗性はすべての植物が保持する基本的な抵抗性と考えることができます。この基礎的抵抗性においては、病原菌の感染に対する抵抗性反応として、抵抗性タンパク質(注5:PRタンパク質)の生産やファイトアレキシン(注6)と総称される低分子抗菌性化合物の生産などが誘導されます。このような植物の病害抵抗性反応においては、MAPキナーゼカスケードと呼ばれるリン酸化シグナル伝達と、続く病害抵抗性関連遺伝子の発現に必要な転写因子が重要な役割を果たしています。これまでの研究で、イネの基礎的抵抗性発現において、OsMKK4-OsMPK3/OsMPK6によるリン酸化カスケードがファイトアレキシン生産を含む抵抗性発現で重要な役割を果たすことや、イネの病害抵抗性に関与するWRKY型転写因子(注7)のひとつであるOsWRKY53により病害抵抗性関連遺伝子の発現誘導が促され、その過剰発現によりイネの耐病性が増強されることが示されていました。
今回、同研究グループは、これまで不明であった、イネがいもち菌に対する抵抗性を発揮する際に必要なOsMKK4-OsMPK3/OsMPK6のリン酸化カスケードとOsWRKY53の関係を明らかにしました。イネを含む複数の植物種において、WRKY型転写因子がリン酸化を受けることで、そのDNA結合能や転写活性化能の向上が認められていました。そこで、まずOsWRKY53のリン酸化について検討し、OsWRKY53のN末端側に6カ所存在するリン酸化部位が、OsMKK4-OsMPK3/OsMPK6によって試験管内でリン酸化されることを示しました。次に、このリン酸化によって、OsWRKY53のDNA結合能には変化がないものの、イネ細胞内における転写活性化能が顕著に増加することを明らかにしました。MAPキナーゼによりリン酸化されるアミノ酸のセリンを、アスパラギン酸などの負電荷を持つ酸性アミノ酸に置換すると、リン酸基を保持する場合と同様にタンパク質の活性化が起こることが知られています。OsWRKY53のリン酸化部位のアミノ酸であるセリンをアスパラギン酸に置換した疑似リン酸化OsWRKY53を作製したところ、MAPキナーゼによる活性化と同等レベルの転写活性化能を発揮することを見出しました。そこで、この疑似リン酸化OsWRKY53を過剰発現するイネを作出したところ、通常のOsWRKY53を過剰発現させたイネと比較して、いもち病菌に対する耐病性がさらに増強されることが明らかになりました。このような耐病性の向上したイネにおいては、キチナーゼやグルカナーゼに代表されるPRタンパク質や、イネの主要ファイトアレキシンであるモミラクトンの生合成遺伝子など、多くの抵抗性関連遺伝子がいもち菌の感染前から発現していることも明らかになりました(図1)。
これらの結果から、いもち菌感染時のイネの病害抵抗性発現においては、OsMKK4-OsMPK3/OsMPK6によりリン酸化され転写活性化されたOsWRKY53が、種々の抵抗性関連遺伝子の転写を誘導することで、耐病性の発揮につながることが強く示唆されました。また、疑似リン酸化型のOsWRKY53をイネに発現させることで、おそって来るいもち菌の攻撃に対してあらかじめ準備しておく“プライミング効果”が起こり、イネの耐病性向上につながったものと考えられます。本成果は、転写因子OsWRKY53を利用した新たな病害抵抗性イネの育種技術に発展することが見込まれるとともに、イネがどのように基礎抵抗性を駆動しているのか、その分子メカニズムを紐解く大きな足がかりになることが期待されます。
発表雑誌
- 雑誌名
- 「PLOS ONE」 June, 3 , 2014」(オンライン版:6月3日)
- 論文タイトル
- Overexpression of Phosphomimic Mutated OsWRKY53 Leads to Enhanced Blast Resistance in Rice
- 著者
- Tetsuya Chujo, Koji Miyamoto, Satoshi Ogawa, Yuka Masuda, Takafumi Shimizu, Mitsuko Kishi-Kaboshi, Akira Takahashi, Yoko Nishizawa, Eiichi Minami, Hideaki Nojiri, Hisakazu Yamane, Kazunori Okada
- DOI番号
- 10.1371/journal.pone.0098737
- アブストラクト
- http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0098737
問い合わせ先
東京大学大学生物生産工学研究センター 環境保全工学研究室
准教授 岡田 憲典
Tel:03-5841-3070
Fax:03-5841-3070
研究室URL:http://park.itc.u-tokyo.ac.jp/biotec-res-ctr/kampo/
用語解説
- 注1 いもち病
- イネいもち菌とよばれるカビの一種により発生する、イネの主要な病気。感染したイネは大幅に収穫が減少し、味も悪くなってしまう。
- 注2 MAPキナーゼカスケード
- MAP キナーゼ(mitogen-activated protein kinase, 以下MAPK)は、酵母から植物・高等動物に至るまで広く真核生物に保存されたセリン/スレオニンキナーゼで、活性化にともなって核内へと移行し、転写因子との相互作用によるリン酸化を経て転写の制御を行っている。その活性化にはMAPKキナーゼ、さらにMAPKキナーゼキナーゼが必要であり、「カスケード(滝の流れの意)」のようなシグナル伝達を構成することからMAPキナーゼカスケードと呼ばれる。
- 注3 MAMPs PAMPs
- Microbe/Pathogen Associated Molecular Patterns略称で日本語訳では病原体関連分子パターンと呼ばれる。 植物が微生物由来の分子を認識して防御応答を活性化する機構において、植物に防御応答を促す分子群の総称。代表的なものにキチンオリゴ糖、フラジェリンタンパク質、リポ多糖などが知られる。
- 注4 エリシター
- 病原菌に感染した植物が分泌する生体防御反応を誘導する物質の総称。病原菌由来の多糖・オリゴ糖、タンパク質・ペプチドなどがある。
- 注5 PRタンパク質(pathogenesis-related proteins)
- 植物が病原菌に感染した際に生産するタンパク質で、抗菌作用があるものが多く生体防御に使われる。病原微生物由来のMAMPsを感知した植物において産生され健全な植物では見られない。代表的なものにキチナーゼやグルカナーゼ、パーオキシダーゼやオスモチンなどが含まれる。
- 注6 ファイトアレキシン
- 健全な植物には存在せず、病原菌の感染やUV照射、重金属ストレスなどにより誘導的に生産される抗菌性二次代謝産物の総称。イネにはフラボノイドタイプのサクラネチンと、4種の環状ジテルペンタイプ、モミラクトン類・ファイトカサン類・オリザレキシン類・オリザレキシンS、カスベンタイプのオキソデプレッシンの存在が知られている。
- 注7 WRKY型転写因子
- WRKY型転写因子は、MAMPsにより発現誘導を受けるエリシター応答性遺伝子のエリシター応答配列(W-box)に特異的に結合する植物特有の転写因子。ジャスモン酸やサリチル酸応答を示す防御関連遺伝子の発現制御に重要な役割を果たす転写因子として注目されている転写因子ファミリーの一つである。