脂質含有量が多い海洋性微細藻類、円石藻の形質転換に成功 - 脂質合成能力の強化によるバイオ燃料生産に期待 -
- 発表者
-
宇治 利樹(北海道大学大学院水産科学研究院 助教)
井上 広滋(東京大学大気海洋研究所 教授)
遠藤 博寿(科学技術振興機構 さきがけ研究者 兼 東京大学大学院農学生命科学研究科 農学共同研究員)
発表のポイント
◆バイオ燃料や原油代替原料の生産への利用可能性を持つ円石藻は、石灰化した細胞壁を持つため遺伝子導入技術注1)が未確立な海洋性微細藻類で、高度な研究が困難であった。
◆円石藻の中でも脂質を多く含む種を使い、細胞に傷をつけずに細胞壁を除去する手法を開発し、導入した遺伝子が次世代に受け継がれる形質転換技術注2)を確立した。
◆有用脂質合成能力の強化や、二酸化炭素固定メカニズムの解明への貢献が期待される。
発表概要
JST 戦略的創造研究推進事業において、JSTの遠藤 博寿 さきがけ研究者(兼 東京大学 大学院農学生命科学研究科 農学共同研究員)、東京大学 大気海洋研究所の井上 広滋 教授、および北海道大学の宇治 利樹 助教らの研究グループは、海洋性の単細胞藻類である円石藻の遺伝子導入技術、中でも非常に難易度の高い形質転換技術の開発に世界で初めて成功しました。
円石藻には、脂質を著しく多く貯蔵する種や、原油の原料となる脂質を生産する種があり、海洋環境学のみならずバイオエネルギー研究の観点からも注目されています。また円石藻は、その細胞壁上にココリス(円石)と呼ばれる炭酸カルシウムの微細構造を持ち、このココリス形成と光合成を介して海洋の炭素循環に大きな影響を及ぼす重要な生物種でもあります。しかし、円石藻では、高度かつ先進的な研究を進める上で不可欠な遺伝子導入の技術が確立されておらず、長年その開発が望まれていました。
遺伝子導入の障害となっていたのは、円石藻の石灰化した強固な細胞壁でした。本研究グループは、細胞をほとんど傷つけずに細胞壁だけを完全に除去する手法を確立し、細胞の活発な分裂能を保ったまま増殖させることに成功しました。次に、細胞壁を除去した細胞にポリエチレングリコール溶液を用いて外来遺伝子注3)を導入し、その遺伝子が短時間でも細胞内で発現する手法を確立しました。これらの手法を用いて、外来遺伝子を次世代以降でも発現する状態の細胞を作製し、円石藻の形質転換技術の確立に成功しました。
本研究成果は、バイオ燃料や原油代替原料の生産への利用が期待される円石藻の有用脂質合成能力の強化や、ココリス形成時に二酸化炭素を強力に固定するメカニズムの解明の研究を進展させ、エネルギーや環境問題の解決に貢献すると期待されます。
本研究成果は、2016年3月7日(英国時間)に英国科学誌「Scientific Reports」のオンライン速報版で公開されます。
本成果は、以下の事業・研究領域・研究課題によって得られました。
戦略的創造研究推進事業 個人型研究(さきがけ)
研究領域 :「藻類・水圏微生物の機能解明と制御によるバイオエネルギー創成のための基盤技術の創出」
(研究総括:松永 是 東京農工大学 学長)
研究課題 :高脂質含有円石藻の形質転換技術の確立と有用脂質高生産に向けた応用
研究者 :遠藤 博寿(JST さきがけ研究者)
研究実施場所:東京大学大気海洋研究所
研究期間 :平成24年10月~平成28年3月
発表内容
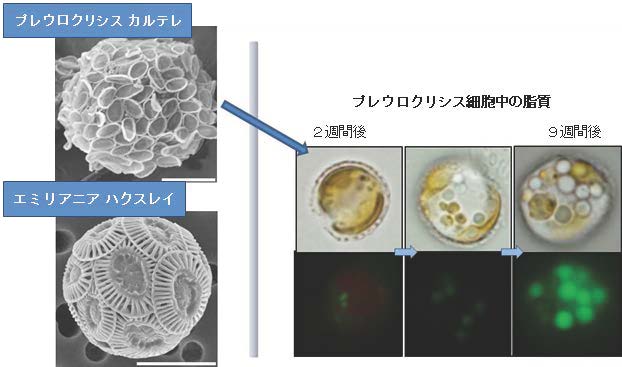
図1 円石藻と脂質の蓄積
左上図:プレウロクリシス カルテレ。本研究の研究対象であり、脂質を多く含み、大量培養が可能な円石藻。
左下図:エミリアニア ハクスレイ。化石燃料に含まれる特殊な脂質を現在も生産し続けている円石藻(画像提供:筑波大学 白岩 善博 教授、猿渡 和子 博士)。細胞表面上に見える楕円状の構造がココリス(炭酸カルシウムの微細構造)。
右 図:プレウロクリシス カルテレの培養開始2~9週間後の細胞内で脂質が蓄積される様子。右下の写真で緑色に見える粒状のものが油滴。(拡大画像↗)
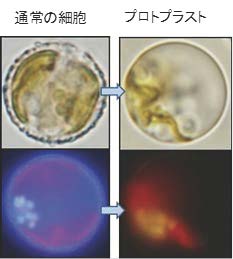
図2 円石藻プレウロクリシス カルテレのプロトプラスト化
細胞壁を持つ細胞(左)と、細胞壁を除去したプロトプラスト(右)。青白く光っているのが細胞壁(左下)。たんぱく質分解および低浸透圧処理により作製したプロトプラストでは、青白い蛍光が消失している(右下)。(拡大画像↗)
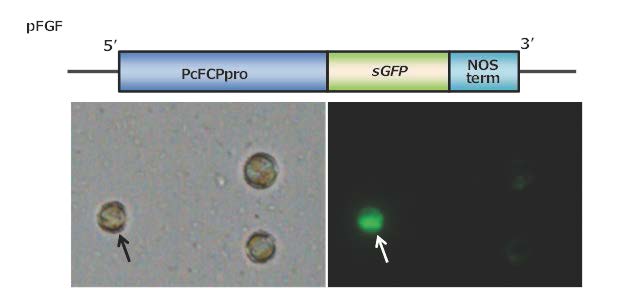
図3 GFP(緑色蛍光たんぱく質)の一過性発現
上図:一過性発現用発現コンストラクト(pFGF)の模式図。
発現させたい遺伝子であるGFP遺伝子の上流と下流それぞれに発現開始調節領域、発現停止調節領域を組み込んだものを作製した。
下図:GFPが発現している細胞(矢印)。
3つの細胞の中で、左の細胞でGFP遺伝子が発現し、緑色の蛍光を放っている。(拡大画像↗)
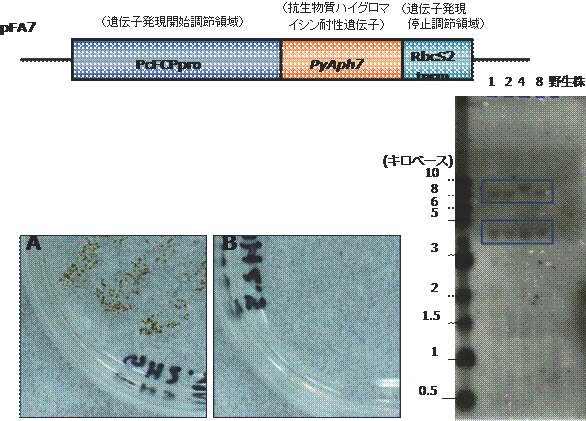
図4 円石藻の形質転換実験
上 図:形質転換用発現コンストラクト(pFA7)の模式図。
発現させたい遺伝子、抗生物質ハイグロマイシン耐性遺伝子の上流と下流それぞれに発現開始調節領域、発現停止調節領域を組み込んだものを作製した。プロモーターは、GFPの一過性発現に成功した時と同じものを選択した。
下左図:抗生物質を加えたプレート培地における培養結果。Aは薬剤耐性遺伝子を導入して形質転換を行った細胞で、Bは野生株。Aでは細胞が増殖している。
下右図:ノザン解析注6)による抗生物質ハイグロマイシン耐性遺伝子のゲノムへの挿入の確認。野生株を除く全ての変異株(No.1、2、4、8)において外来遺伝子の挿入を示すシグナルが確認できる(枠内)。(拡大画像↗)
<研究の背景と経緯>
微細藻類は、サトウキビやトウモロコシなどの穀物を原材料とするバイオ燃料生産とは異なり、食糧生産と競合しないため、次世代のバイオ燃料の有力候補として期待されています。海洋性の微細藻類である円石藻の中には、細胞内に油滴を形成して脂質を著しく多く蓄積する種や、原油・天然ガスの起源生物の1つと考えられている種があり、非常に興味深い生物群です(図1)。また円石藻は、細胞表面に炭酸カルシウムからなるココリス (cocco=円、lith=石)と呼ばれる微細構造を形成することで、二酸化炭素を大量に固定し、このココリス形成と光合成を通じて海洋の炭素循環に寄与する重要な生物でもあります(図1)。近年では円石藻の光学特性注4)の研究が進められ、新たなナノデバイス開発の観点からも、円石藻の研究の重要性が認識されています。
しかし、脂質の合成や炭酸カルシウム形成のメカニズムの詳細は、特に遺伝子レベルではほとんど研究されてきませんでした。これは、遺伝子の過剰発現や遺伝子ノックアウトといった高度な研究技術が確立されていないことが大きな要因として挙げられます。そこで、本研究グループは、円石藻の中でも特に脂質を多く含むプレウロクリシス カルテレ(Pleurochrysis carterae)という種を用いて形質転換技術の確立に取り組みました。
<研究の内容>
円石藻は、石灰化した非常に強固な細胞壁を持つため、従来の方法では外来遺伝子を導入することは不可能でした。この問題を解決するために、本研究グループは、たんぱく質分解酵素と低浸透圧溶液を併用することで細胞に吸水を促し、あたかもブドウの中身だけを皮から取り出すように、石灰化した細胞壁から脱皮させる形で完全に除去する手法を開発しました(図2)。
さらに、この手法でほとんど無傷のままプロトプラスト注5)化した細胞に、ポリエチレングリコール溶液を用いて外来遺伝子を導入する手法も確立し、外来遺伝子を円石藻の細胞内で発現させることに世界で初めて成功しました。図3に示した写真が、GFP(緑色蛍光たんぱく質)遺伝子を導入し、その遺伝子が細胞中で発現して緑色蛍光を発している様子です。
外来遺伝子の発現には、ごく短期間で終わってしまうもの(一過性発現)と、恒常的に、つまり世代を越えて常に発現するもの(恒常的発現)があります。後者 は「形質転換」と呼ばれ、生物があらかじめ持っているゲノムの中に外来遺伝子を組み込むことが必須ですが、一般に強固な細胞壁を持ち、プロトプラスト化が難しい海洋性の藻類では非常に難易度の高い技術です。しかし、本研究グループが開発した技術を用いることで、外来遺伝子をゲノムに組み込み、次世代以降でも発現する状態の細胞を作り出すことに成功しました。図4に示す写真は、円石藻にある種の薬剤の耐性遺伝子を導入し、薬剤耐性が次の世代にも受け継がれ、細胞が増殖している様子です。
現在行われている形質転換実験の多くは高価な装置が必要ですが、本技術は簡単に入手可能な試薬のみで実験できます。そのため、形質転換技術が未確立である、他の有用な藻類における技術開発の研究にも寄与することが期待されます。
<今後の展開>
本技術により、円石藻が本来持たない遺伝子を細胞内で発現させることが可能になりました。バイオ燃料や有用物質としての利用に適した脂質の合成能力を強化するための研究が大きく前進することが期待されます。現在、バイオ燃料生産の候補とされる微細藻類のほとんどは淡水性ですが、円石藻は海洋性なので、培養のための真水を作るエネルギーやコストが不要で、バイオ燃料や有用物質を効率よく生産できる可能性があります。また、ココリス形成時の強力な二酸化炭素固定メカニズムについても研究が進むことにより、海洋の酸性化問題解決の一助となる知見が得られることが期待されます。
発表雑誌
- 雑誌名
- 「Scientific Reports」
- 論文タイトル
- “Stable Nuclear Transformation System for the Coccolithophorid Alga Pleurochrysis carterae.”
(円石藻プレウロクリシス カルテレの核の形質転換系) - 著者
- DOI番号
- 論文URL
問い合わせ先
科学技術振興機構 さきがけ研究者
(兼 東京大学 大学院農学生命科学研究科)
農学共同研究員 遠藤 博寿(エンドウ ヒロトシ)
Tel:04-7136-6215
Fax:04-7136-6216
E-mail:aenhiro@mail.ecc.u-tokyo.ac.jp
東京大学大気海洋研究所
教授 井上 広滋(イノウエ コウジ)
Tel:04-7136-6212
Fax:04-7136-6216
研究室URL:http://darwin.aori.u-tokyo.ac.jp/inouelab/index.html
用語解説
- 注1 遺伝子導入技術
- 外来遺伝子(注3を参照)を細胞内に導入し、発現させる技術。
- 注2 形質転換技術
- 遺伝子導入技術の1つで、導入した遺伝子が次世代に受け継がれ、かつ恒常的に発現する状態にする技術。
- 注3 外来遺伝子
- その生物が本来持たない遺伝子。本研究では、GFP(緑色蛍光たんぱく質/クラゲ由来)や、Aph7(ハイグロマイシン耐性遺伝子/バクテリア由来)などが、その利用例にあたる。
- 注4 円石藻の光学特性
- ココリスに光が当たる方向により、光散乱がどのように異なるかの特性。近年、細胞から分離した状態の微細な結晶の方向を永久磁石程度の磁場を利用して非接触で動かす技術を開発した成果などが報告されており、研究が大きく前進している。
Miyashita,Y. Iwasaka, M. and Endo, H. J. Appl. Phys., 117, 17E130-1-17E130-4 (2015) Mizukawa, Y., Miyashita, Y., Satoh, M., Shiraiwa S. & Iwasaka, M. Sci. Rep., doi 10.1038/srep13577 (2015) - 注5 プロトプラスト
- 多くの植物や藻類が細胞の最外層に持つ細胞壁を欠いた細胞。多くの藻類は細胞壁を欠いた状態にすることにより、遺伝子導入の成功率が高まる。
- 注6 ノザン解析
- 膜上に張り付けたDNAから、特定の配列の有無を調べる実験。