植物の栄養による生長制御のためのネットワークにおける硝酸イオン-CPK-NLPシグナル伝達の発見
- 発表者
-
Kun-hsiang Liu(ハーバード大学医学系大学院)
Yajie Niu(ハーバード大学医学系大学院)
小西 美稲子(東京大学生物生産工学研究センター 特任助教)
Yue Wu (ハーバード大学医学系大学院)
Hao Du(ハーバード大学医学系大学院)
Hoo Sun Chung(ハーバード大学医学系大学院)
Lei Li, Marie Boudsocq(ハーバード大学医学系大学院)
Matthew McCormack(ハーバード大学医学系大学院)
前川 修吾(東京大学生物生産工学研究センター 日本学術振興会特別研究員)
石田 哲也(東京大学生物生産工学研究センター 特任助教 当時)
Chao Zhang (カリフォルニア大学サンフランシスコ校)
Kevan Shokat(カリフォルニア大学サンフランシスコ校 教授)
柳澤 修一(東京大学生物生産工学研究センター 教授)
Jen Sheen (ハーバード大学医学系大学院 教授)
発表のポイント
◆硝酸態の窒素栄養(硝酸イオン)の吸収に伴って起こる栄養応答関連遺伝子の発現の分子メカニズムを明らかにしました。
◆栄養シグナルと共役したCa2+シグナル伝達ネットワークがトランスクリプトーム(注1)、代謝、地上部-根のバランス調整など器官形成上の可塑性を統合していることを明らかにしました。
◆植物の栄養利用効率を向上させる技術開発への貢献が期待されます。
発表概要
栄養シグナル伝達は遺伝子発現と代謝、生長を統合し調和させています。植物の窒素源である硝酸イオンはシグナル分子としての作用も持ち、遺伝子の発現を制御して植物の代謝や生長を調節しています。シロイヌナズナの葉と根の両方で硝酸イオンの吸収に伴って細胞内Ca2+濃度が上昇すること、これによりCa2+依存性タンパク質リン酸化酵素(注2)であるCPK10、CPK30、CPK32が活性化され、硝酸シグナルに応答した遺伝子発現を司るNLP転写因子(注3)をリン酸化することを発見しました。リン酸化されたNLPは活性型となり、窒素同化・輸送を担う遺伝子を含む様々な遺伝子の発現を引き起こします。cpk10 cpk30 cpk32三重変異体とnlp7変異体で同様に硝酸イオンによって誘導される地上部の生育と根系の確立のシステム全体に支障が観察され、栄養シグナルと結びついたCa2+シグナル伝達ネットワークはトランスクリプトーム、細胞の代謝、地上部-根のバランス調整など器官形成上の可塑性を統合していることが示されました。
発表内容
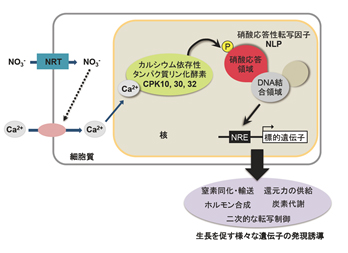
図1 硝酸シグナルに応答した遺伝子発現制御が植物の窒素利用と成長を促すモデル図
(拡大画像↗)
植物は土壌中の窒素を吸収・同化し、タンパク質や核酸など窒素原子を含むさまざまな生体分子を生合成して生長しています。土壌中の窒素含量は高い農業生産を得るためには十分ではないため、近代農業では大量の窒素肥料の施肥を行なっています。しかしながら、大量の窒素肥料の使用は、河川や海の富栄養化を引き起こし、酸性雨の元凶ともなり、ハーバー・ボッシュ法を用いた窒素肥料の化学合成過程においてCO2が排出されるなど、環境への大きな負荷を与えています。農業生産性の維持・向上と環境保全の両立させるためには植物の窒素利用効率を高めることが重要となっています。
植物の主たる窒素源である硝酸イオンは、窒素を効率よく吸収して活用するためのシステムを制御するシグナル分子としての役割を担っています。硝酸シグナルは、窒素同化に関わる遺伝子や窒素吸収と利用の調節に関わる遺伝子の発現を変化させて、窒素利用システムを直接的に調節しています。また、窒素の利用に関連する遺伝子以外にも、炭素や植物ホルモンの代謝、ミネラルの輸送や転写調節に関わる遺伝子など多数の遺伝子の発現も調節していることが知られており、硝酸シグナルによる制御は植物の窒素利用とそれに応じた代謝・成長調節の鍵を握ると考えられてきました。 この硝酸シグナルによって引き起こされる遺伝子発現は、NLP転写因子が担うことが数年前に明らかにされ、また、NLP転写因子は硝酸シグナルによって活性化されることが明らかにされていましたが、活性型NLPの実体や硝酸シグナルによるNLPの活性化メカニズムは不明なままでした。
今回、超高感度バイオセンサーのライブイメージングにより、シロイヌナズナの葉と根において硝酸イオンの取り込みに伴ってCa2+シグナル伝達が起こること、このシグナル伝達によってCa2+依存性タンパク質リン酸化酵素であるCPK10、CPK30、CPK32が活性化され、これらによってNLP転写因子がリン酸化されることを発見しました。リン酸化されたNLP転写因子が活性型であり、同化・輸送を担う遺伝子のみならず、炭素や植物ホルモンの代謝経路や転写制御因子の遺伝子などの発現を促進するという制御メカニズムが明らかとなりました。
また、cpk10 cpk30 cpk32三重変異体にCPK10(M141G)改変タンパク質を導入することにより胚性致死を回避して行った解析から、cpk10 cpk30 cpk32変異体とnlp7変異体のいずれもが同様に硝酸シグナルによって誘導される地上部の生育と根系の確立に支障をきたしていることが確認され、硝酸イオン-CPK-NLPシグナル伝達経路が、トランスクリプトーム、細胞の代謝、地上部-根のバランス調整など器官形成上の可塑性を統合しており、植物の生長制御ネットワークで中心的な役割を担っていることが明らかとなりました。
この統合的な遺伝子発現機構により、窒素同化とそれに応じた代謝・成長の調節が実現していると考えられます。NLP転写因子は植物種を問わず保存されていることから、この発見は作物の窒素利用効率の向上とバイオマス増産のための技術開発に貢献することが期待されます。
発表雑誌
- 雑誌名
- :Nature Vol. 545, 311-316 (オンライン版:5月10日)
- 論文タイトル
- :Discovery of nitrate–CPK–NLP signaling in central nutrient–growth networks
- 著者
- :Kun-hsiang Liu*, Yajie Niu, Mineko Konishi, Yue Wu, Hao Du, Hoo Sun Chung, Lei Li, Marie Boudsocq, Matthew McCormack, Shugo Maekawa, Tetsuya Ishida, Chao Zhang, Kevan Shokat, Shuichi Yanagisawa*, Jen Sheen* (*共同責任著者)
- DOI番号
- :10.1038/nature22077
- 論文URL
- :https://www.nature.com/nature/journal/vaop/ncurrent/full/nature22077.htm
問い合わせ先
- 東京大学生物生産工学研究センター 植物機能工学部門
教授 柳澤 修一(ヤナギサワ シュウイチ) - Tel:03-5841-5258
Fax:03-5841-8030 - Email: asyanagi<at>mail.ecc.u-tokyo.ac.jp <at>を@に変えてください。
研究室URL:http://park.itc.u-tokyo.ac.jp/ppk/home/
用語解説
- 注1 トランスクリプトーム
- 細胞内の全転写産物(mRNA)。その細胞または組織における遺伝子発現全体を反映している。
- 注2 Ca2+依存性タンパク質リン酸化酵素
- タンパク質にリン酸基を付加する酵素だが、Ca2+と結合したときにのみその触媒機能を発揮する性質を持つため、細胞内Ca2+のセンサーとしてCa2+シグナルの伝達を担っている。
- 注3 NLP転写因子
- 植物が持つ、遺伝子発現の調節を担うタンパク質の一つ。それぞれの植物種が複数のNLPタンパク質を持っている。ゲノムDNA上の特定の塩基配列に結合して、近傍の遺伝子の発現量を増加させる。NLPタンパク質は、硝酸シグナルが伝達されたときのみ遺伝子発現を促進し、硝酸シグナルが伝達されない場合は機能しないという特徴を持つ。