植物に乾燥・高温耐性を付与する転写因子DREB2Aが活性化する仕組みを解明
-干ばつや高温に強い作物の開発に期待-
- 発表者
-
森本 恭子(東京大学大学院農学生命科学研究科 応用生命化学専攻 特任研究員;当時)
篠崎 和子(東京大学大学院農学生命科学研究科 応用生命化学専攻 教授)
大濱 直彦(東京大学大学院農学生命科学研究科 応用生命化学専攻 特任助教;当時)
城所 聡(東京大学大学院農学生命科学研究科 応用生命化学専攻 助教)
溝井 順哉(東京大学大学院農学生命科学研究科 応用生命化学専攻 講師)
高橋 史憲(理化学研究所 環境資源科学研究センター 研究員)
戸高 大輔(東京大学大学院農学生命科学研究科 応用生命化学専攻 特任助教)
最上 惇郎(東京大学大学院農学生命科学研究科 応用生命化学専攻 特任助教)
佐藤 輝(東京大学大学院農学生命科学研究科 応用生命化学専攻 博士課程学生;当時)
秦 峰(国際農林水産業研究センター 研究員;当時)
金 俊植(理化学研究所 環境資源科学研究センター 基礎科学特別研究員)
深尾 陽一朗(奈良先端科学技術大学院大学 特任准教授;当時)
藤原 正幸(奈良先端科学技術大学院大学 助教;当時)
篠崎 一雄(理化学研究所 環境資源科学研究センター センター長)
発表のポイント
◆植物の干ばつと高温の両方の耐性獲得に働く転写因子(注1)DREB2Aは、通常条件下では植物細胞内で速やかに分解されますが、この分解はBPM-CUL3 E3リガーゼ(注2)によって引き起こされることを明らかにしました。
◆BPM-CUL3 E3リガーゼによる分解から逃れることが、干ばつ・高温時におけるDREB2A活性化の正体であることが示され、10年以上未解明であったDREB2A活性化の仕組みが分子レベルで明らかになりました。
◆高温と干ばつが同時に起こると作物は大きな被害を受けますが、両方の耐性獲得に働くDREB2Aの活性を制御することで作物の減収を防ぐ技術が開発されると期待されます。
発表概要
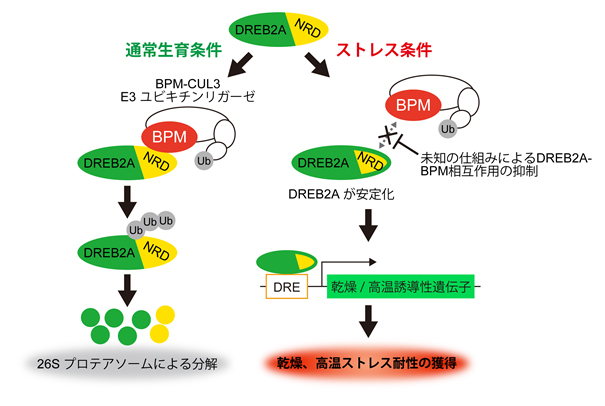
図1 植物の干ばつと高温の両方の耐性獲得に働く転写因子DREB2Aの構造中のNRDは、タンパク質の分解系であるBPM-CUL3 E3リガーゼが認識して結合する配列である。通常生育条件下では、その結合によりDREB2Aはユビキチン化されプロテアソーム系に運ばれ分解される。乾燥・高温ストレス時にはリン酸化などによりNRDが修飾されたり、または未知のタンパク質によってNRDがマスクされたりして、BPM-CUL3 E3リガーゼによる分解系から逃れることで活性化する。活性化したDREB2Aは耐性遺伝子の発現を誘導し乾燥や高温耐性が獲得される。(拡大画像↗)
東京大学と理化学研究所及び国際農林水産業研究センターの共同研究グループは、植物の干ばつと高温の両方の耐性獲得に働く転写因子DREB2Aの活性化の機構の解明に成功しました。植物は、高温や乾燥などの環境ストレスを受けると、多数の耐性遺伝子の発現を誘導することにより耐性を獲得します。干ばつと高温の複合ストレス下では、植物は蒸散による体温の低下の仕組みを働かせることができなくなることから、大きなダメージを受けます。DREB2Aは高温と乾燥の両方のストレスに応答して活性化することから、この複合ストレス時に重要な機能を示します。DREB2Aはそのままでは活性を示しませんが、DREB2Aの構造からNRDと名付けられた中央部のアミノ酸配列を除くと、安定な活性型(DREB2A CA)に変換され、これを植物に導入すると乾燥と高温の両方のストレスに対する耐性が向上します。しかし、このDREB2A CAの活性化の分子機構は、10年以上不明なままでした。本研究によって、NRDはタンパク質の分解系であるBPM-CUL3 E3リガーゼが認識して結合する配列であり、その結合によりDREB2Aの分解機構が働くことが明らかになりました(図1)。DREB2A CAはNRDを持たないため、BPM-CUL3 E3リガーゼによって認識されず分解されないため安定化します。このように、NRDを除くとDREB2A CAが安定化して活性型に変換される機構が解明されました。植物中ではストレス時特異的にリン酸化などの修飾や未知のタンパク質によってNRDがマスクされる機構が考えられます(図1)。この研究により、植物の高温や乾燥ストレスに対する耐性獲得機構の理解が進むとともに、高温や干ばつに対する耐性を向上させた作物の開発への応用が期待されます。
発表内容
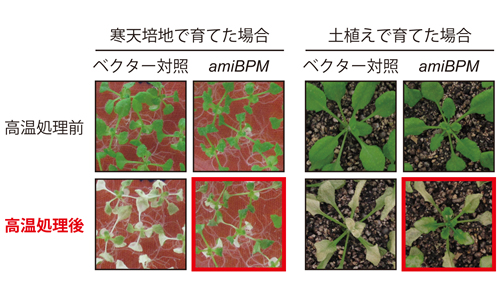
図2 新たに同定された、CUL3 E3リガーゼの基質アダプターであるBPMの働きを抑えた植物(amiBPM)では、高温耐性の向上が示された。 (拡大画像↗)
地球レベルの環境劣化が進み、異常気象による農業被害が深刻化しています。さらに地球温暖化の影響によって、急激な温度変化や干ばつなどが頻発するようになると予測されています。このため、環境ストレスに高い耐性を持つ作物の育種が重要となっています。植物は、環境ストレスを受けると、さまざまな遺伝子の発現を変化させることにより耐性を獲得します。DREB2Aは、高温や乾燥の複合ストレスに対する植物の耐性獲得機構において中心的な役割を果たす転写因子で、植物が乾燥と高温の両方のストレスに対する耐性を獲得するために機能する多数の遺伝子群の働きを活性化させることができます。このため、環境ストレス耐性作物の分子育種における有望な因子と考えられています。しかし、DREB2Aをそのまま植物中で強く発現させても働きを示しません。共同研究グループは10年前、DREB2Aの構造からNegative regulatory domain (NRD)と名付けた30アミノ酸の配列を取り除くと活性型(DREB2A CA)に変換できることを発見しました。さらに、DREB2A CAを植物中で高発現すると、多数の標的遺伝子の発現が強く誘導されて、乾燥と高温ストレスに対する耐性が向上することも示しました。しかし、なぜNRD配列を除くとDREB2Aは活性型に変換されるのか、その分子メカニズムは長い間不明のままでした。
今回、本共同研究グループは、シロイヌナズナを用いてDREB2Aのタンパク質複合体を精製し、質量分析装置を用いることでDREB2Aと相互作用するタンパク質を解析しました。その結果、DREB2A相互作用タンパク質として、CUL3 E3リガーゼの基質アダプターであるBPMを同定しました。BPMはDREB2Aと転写因子が機能する細胞の核内で相互作用します。この時、DREB2AのNRD配列が特異的にBPMと相互作用することを見出しました。このBPMの働きを抑えた植物では、高温および乾燥ストレス条件下でDREB2Aタンパク質が蓄積し、DREB2Aの標的遺伝子群の誘導が増加していることが示されました。その結果として、この植物では高温耐性が向上しました(図2)。このようにDREB2Aの相互作用因子としてBPM-CUL3 E3リガーゼを同定したことにより、長い間追い求めていたDREB2A CAの活性型への変換の原因が明らかになりました。
高温などのストレス時の植物中では、リン酸化などのNRDの修飾や未知のタンパク質によるNRDのマスクにより、BPM-CUL3 E3リガーゼがDREB2Aを認識できなくなると推測されます(図1)。本研究では、質量分析装置を用いてDREB2Aタンパク複合体を分析することで、BPM-CUL3 E3リガーゼとDREB2Aの関係を明らかにすることに成功しました。さらに、様々な転写因子においてもNRDに類似した配列が見出されることから、代謝回転が早い転写因子の分解制御においてBPM-CUL3 E3リガーゼが重要な役割を果たしていることが考えられました。
以上の結果より、これまで10年以上の長い間不明のままであったDREB2A CAの活性化の機構が明らかになりました。今回、DREB2Aの相互作用因子として単離されたBPM-CUL3 E3リガーゼは、DREB2Aタンパク質をユビキチン化(注3)してプロテアソーム分解系に運ぶシステムであり、NRD配列を特異的に認識して結合しDREB2Aを分解系に運びます。NRDを持たないDREB2A CAは、分解系から逃れ安定化することで活性化することが示されました。DREB2Aは多くの耐性遺伝子群を制御して乾燥と高温の両方のストレス耐性を向上しますが、成長には負の影響を示すことが明らかにされています。そのため、不必要な時はBPM-CUL3 E3リガーゼによりユビキチン化してタンパク質の分解系に運んでいると考えられました。これらのシステムを効率よく制御することで、高温や干ばつに対する耐性を向上させた作物の開発への応用が期待されます。
発表雑誌
- 雑誌名
- :Proc. Natl. Acad. Sci. USA
- 論文タイトル
- :BPM-CUL3 E3 ligase modulates thermotolerance by facilitating negative regulatory domain-mediated degradation of DREB2A in Arabidopsis
- 著者
- :#Kyoko Morimoto, #Naohiko Ohama, Satoshi Kidokoro, Junya Mizoi, Fuminori Takahashi, Daisuke Todaka, Junro Mogami, Hikaru Sato, Feng Qin, June-Sik Kim, Yoichiro Fukao, Masayuki Fujiwara, Kazuo Shinozaki, Kazuko Yamaguchi-Shinozaki* #These authors contributed equally to this work.
- DOI番号
- :10.1073/pnas.1704189114
- 論文URL
- :www.pnas.org/cgi/doi/10.1073/pnas.1704189114
問い合わせ先
東京大学大学院農学生命科学研究科応用生命化学専攻 植物分子生理学研究室
教授 篠崎 和子(しのざき かずこ)
Tel:03-5841-8137
Fax:03-5841-8009
E-mail:akys <アット>mail.ecc.u-tokyo.ac.jp <アット>を@に変えてください。
用語解説
- 注1 転写因子
- 標的とする遺伝子のプロモーター配列(DNA)に結合し、その遺伝子の発現を活性化したり抑制したりして調節するDNA結合性タンパク質。DREB2Aは高温及び乾燥ストレス誘導性遺伝子発現の活性化因子として働いている。
- 注2 BPM-CUL3 E3リガーゼ
- CUL3 E3リガーゼは、RING-finger型E3リガーゼであり、E1、E2と共に標的タンパク質のユビキチン化を触媒する。BPMは基質認識サブユニットを形成する。
- 注3 ユビキチン化
- タンパク質修飾の一種で、ユビキチンリガーゼの働きにより基質タンパク質に付加される。多くの場合、ユビキチン自身もさらに重合しポリユビキチン鎖を形成する。ポリユビキチン修飾されたタンパク質は、プロテアソームにより認識され分解される。