東京大学 農学生命科学研究科 研究成果
味覚修飾タンパク質ネオクリンのpH依存的活性を決定づけるアミノ酸残基の同定
- 発表者
- 中島 健一朗 (東京大学大学院農学生命科学研究科 応用生命化学専攻・特任助教(当時))
横山 可那子 (東京農業大学 応用生物科学部・学部生(当時))
小泉 太一 (東京大学大学院農学生命科学研究科 応用生命化学専攻・修士課程)
古泉 文子 (東京大学大学院農学生命科学研究科 応用生命化学専攻・博士課程(当時))
朝倉 富子 (東京大学大学院農学生命科学研究科 応用生命化学専攻・特任准教授)
寺田 透 (東京大学大学院農学生命科学研究科 アグリバイオインフォマティクス教育研究ユニット・特任准教授)
益田 勝吉 (サントリー生物有機科学研究所)
伊藤 圭祐 (東京大学大学院農学生命科学研究科 応用生命化学専攻・特任研究員(当時))
清水(井深) 章子 (東京大学大学院農学生命科学研究科 応用生命化学専攻・特任研究員(当時))
三坂 巧 (東京大学大学院農学生命科学研究科 応用生命化学専攻・准教授)
阿部 啓子 (東京大学大学院農学生命科学研究科 応用生命化学専攻・特任教授)
発表概要
酸っぱいものを甘く感じさせる味覚修飾タンパク質(注1)の一つであるネオクリンは、酸性サブユニット(NAS)と塩基性サブユニット(NBS)のヘテロダイマーで、自身が甘味を持つ上、酸っぱいものを同時に味わうと一層強い甘味を示す。我々はネオクリンの甘味強度の変化に、タンパク質中のたった1つの残基(NBSサブユニットの11番目に位置するヒスチジン)が、あたかも一般酸塩基触媒のように、決定的に寄与していることを明らかにした。またこのアミノ酸残基に様々な点変異を導入することで、pH依存的な活性が大きく変化するタンパク質を作り出すことができた。
発表内容
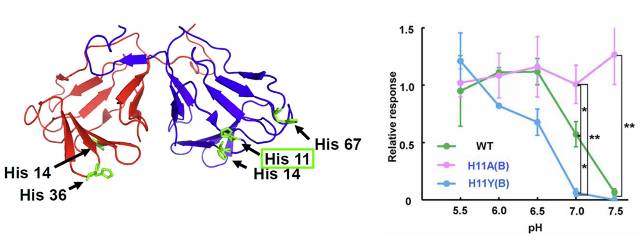
図:ネオクリン変異体における甘味活性
左 ネオクリンにおけるヒスチジン残基の位置。活性に大きな影響を与えるNBSサブユニットの11番目に位置するヒスチジン残基を緑枠で示した。
右 野生型(緑色)、NBSサブユニットの11番目に位置するヒスチジン残基をアラニン(桃色)あるいはチロシン(水色)に変えた変異体におけるpH依存的な甘味活性。
味覚修飾タンパク質を口に含んだ後に、酢酸やクエン酸といった酸味を呈するものを味わうと非常に甘く感じるという現象が知られている。この効果は、一見、味覚修飾タンパク質が「酸味を甘味に変換する」かのように感じてしまう。この活性を持つものとして、熱帯植物の果実中に含まれるネオクリン、ミラクリンの2種のタンパク質の存在が知られている。
味覚修飾タンパク質が酸によって甘味を誘導する機構について、我々は以前に麹菌発現系を用いてネオクリンの変異体を作製し検証を行った (Nakajima, K. et al., FASEB J., 22, 2323-2330 (2008))。ネオクリンのNASにはヒスチジン残基が2つ、NBSにはそれが3つ存在する(図)。これら5つのヒスチジン残基を全てアラニンに置換した変異体(HA変異体)においては野生型ネオクリンと異なり、弱酸性~弱アルカリ性の領域でほぼ一定の甘味を示す甘味タンパク質としての性質を示した。したがってネオクリンがpHに依存して甘味を変化させる過程で、ヒスチジン残基が深く関与していることが示唆された。
今回の報告では、ネオクリンに存在する5つのヒスチジン残基の変異体をデザインするにあたり、1か所ずつをそれぞれアラニンに置換した変異体を作製して甘味強度の測定を行った。5種類の変異体いずれにおいても、弱酸性条件下(pH6.0)においては強い甘味活性を有していた。しかしpH7.5における応答性は変異体間で大きな差があり、野生型と同様にほとんど活性がないものや、HA変異体と同様に強い活性を示すものが認められた。これより5か所のヒスチジン残基が、pH依存的な甘味活性に同等に寄与するわけではないことが判明した。特にNBSサブユニットの11番目に位置するヒスチジンをアラニンに置換した変異体は、5か所を変異させたHA変異体と同様の活性を示したことから、たった1つの残基がpH依存的甘味活性を決定しうることが分かった(図)。
さらにNBSサブユニットの11番目に位置するヒスチジンについて、多種類のアミノ酸に置換した変異体の活性を評価したところ、この残基側鎖に芳香族を有することが味覚修飾活性に必須であることが判明した。さらにフェニルアラニンあるいはチロシンに置換した変異体においては、中性条件における甘味がほとんど消失しており、野生型と異なる味覚修飾活性を有する変異体の作出に成功したといえる(図)。
以上の結果より、ネオクリンがpH依存的に甘味活性を変化させるのに、タンパク質中のたった1つの残基が決定的に寄与していること、またこのアミノ酸残基に様々な点変異を導入することで、pH依存的な活性が大きく変化するタンパク質を作り出すことができた。味覚修飾タンパク質の不思議の解明に、端緒を見出すことができた。
発表雑誌
"Identification and Modulation of the Key Amino Acid Residue Responsible for the pH Sensitivity of Neoculin, a Taste-Modifying Protein"
Ken-ichiro Nakajima, Kanako Yokoyama, Taichi Koizumi, Ayako Koizumi, Tomiko Asakura, Tohru Terada, Katsuyoshi Masuda, Keisuke Ito, Akiko Shimizu-Ibuka, Takumi Misaka, and Keiko Abe
PLoS ONE 6(4): e19448 (2011)
doi:10.1371/journal.pone.0019448
問い合わせ先
応用生命化学専攻 生物機能開発化学研究室 准教授
Tel: 03-5841-8100
E-mail: amisaka@mail.ecc.u-tokyo.ac.jp
ホームページ http://park.itc.u-tokyo.ac.jp/biofunc/
用語解説
味覚修飾タンパク質を摂取した後に、酢酸やクエン酸といった酸味を呈するものを口にすると非常に甘く感じる。この活性を持つものとして、熱帯植物の果実中に含まれるネオクリン、ミラクリンの2種のタンパク質の存在が知られている。