東京大学 農学生命科学研究科 研究成果
植物ホウ素輸送体のmRNAの蓄積はmRNA安定性によって制御されている
- 発表者
- 田中 真幸 (東京大学大学院農学生命科学研究科応用生命化学専攻・特任助教)
高野 順平 (北海道大学大学院農学研究院応用生命科学部門・助教)
千葉 由佳子 (北海道大学創成研究機構・特任助教)
Fabien Lombardo (東京大学大学院農学生命科学研究科応用生命工学専攻・博士研究員;当時)
小笠原 由貴 (東京大学大学院農学生命科学研究科応用生命工学専攻・修士課程;当時)
尾之内 均 (北海道大学大学院農学研究院応用生命科学部門・准教授)
内藤 哲 (北海道大学大学院農学研究院応用生命科学部門・教授)
藤原 徹 (東京大学大学院農学生命科学研究科応用生命化学専攻・教授)
発表概要
シロイヌナズナのホウ素吸収を担うホウ酸チャネルNIP5;1(注1)の転写産物(mRNA)はホウ素欠乏条件で多く蓄積することが知られています。本研究ではホウ素栄養条件に応じたNIP5;1mRNAの蓄積量の変動は、NIP5;1mRNAの分解の制御によって起こっていることを明らかにしました。
発表内容
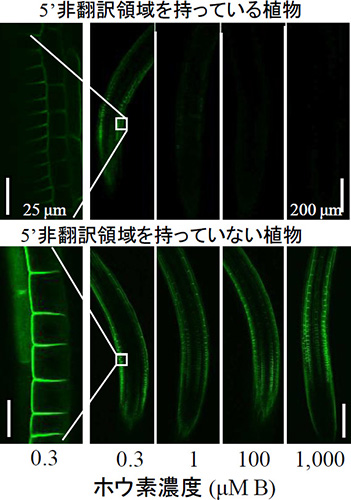
図1 5’非翻訳領域がNIP5;1の高ホウ素濃度に応答した発現に与える影響 (拡大画像↗)
NIP5;1 5’UTRをGFP-NIP5;1遺伝子と連結したもの、あるいは連結しないものをNIP5;1プロモーター制御下で発現させたシロイヌナズナ形質転換体の根のGFP 蛍光を観察した。 NIP5;1 5’UTRを持っていないと、ホウ素過剰条件(1000 μM B)でもGFP蛍光は消えない。
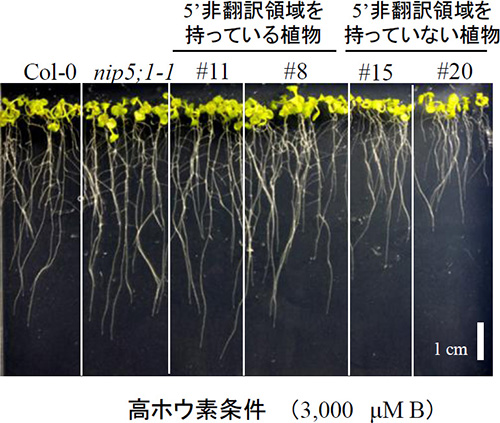
図2 5’非翻訳領域が植物の成長に与える影響 (拡大画像↗)
図1の形質転換植物を用いて、過剰ホウ素条件(3000μM B)における、成長を観察した。NIP5;1 5’UTRを持った形質転換植物は、野生型、NIP5;1の欠損株(nip5;1-1)と比較して、その成長に違いは見られない。一方、 NIP5;1 5’UTRを持たない形質転換植物は、野生型、NIP5;1の欠損株と比較して、明らかに成長が抑制されている
ホウ素は植物の生育に必須な微量元素であり、細胞壁の構造維持に重要な役割を果たしていることが知られています。 私たちは、2006年にシロイヌナズナの根において、ホウ素の吸収に重要なホウ酸チャンネルNIP5;1(注1)を発見しました。NIP5;1はホウ素欠乏条件においてホウ素を土壌から吸収するために必須な遺伝子であり、ホウ素欠乏条件で育てると、ホウ素十分条件で育てた時に比べてNIP5;1のmRNAの蓄積は約10倍上昇することが明らかとなっています。
本研究では、ホウ素に応答したNIP5;1の発現制御機構を明らかにすること目的とし実験を進めました。その結果、ホウ素に応答したNIP5;1のmRNAの蓄積は、mRNAの分解によって制御されていることを発見しました。つまり、ホウ素が十分に存在する条件では、mRNAが素早く分解され、NIP5;1mRNAは蓄積できなく、ホウ素が低濃度に存在する条件では、mRNAは分解されず、NIP5;1mRNAは蓄積します。このmRNAの分解を制御している領域が5’非翻訳領域内(注2)に存在していることも明らかにしました。NIP5;1の5’非翻訳領域を持たない形質転換植物はホウ素過剰条件でも、mRNAが分解されず蓄積し続けるため(図1)、植物はホウ素を過剰に取り込み、結果として成長が抑制されることが示されました(図2)。
植物は動くことができないため、その土地の環境に適応していかなければなりません。土壌中のホウ素濃度は概して均一ではなく、土壌中のホウ素濃度の変化に対応し、植物はホウ素を効率的に吸収、あるいは抑制するように調節しなければなりません。様々な土壌中のホウ素濃度環境に適応するため、mRNA分解による制御が行われていると推察されます。この実験により、植物の栄養輸送体における5’非翻訳領域を介したmRNA分解の制御メカニズムを初めて示しました。この制御機構を応用することで、様々な不良土壌でも効率的にホウ素を吸収、あるいは抑制できる作物を作出することに大きく貢献できると考えています。
発表雑誌
http://www.plantcell.org/content/23/9/3547.abstract
問い合わせ先
用語解説
チャンネル(輸送体)は膜を横切って物質輸送を行うために存在する膜タンパク質。分子またはイオンを通過させるための孔(穴)を形成する。通常イオンや分子の濃度勾配に従い輸送させる。シロイヌナズナのホウ酸チャンネルNIP5;1はホウ酸の輸送体で、主に根の表皮に存在し、土壌中のホウ素を取り込むために重要な役割を果たす。
(注2) 5’非翻訳領域転写開始点から翻訳開始点の間に存在する翻訳されない領域。