発表概要
真核生物のゲノムDNAは細胞の核内にヒストン8量体に結合して存在しています。ヒストン8量体にゲノムDNAは2回弱巻きついており、その構造をヌクレオソームと呼びます。ゲノムワイドなヌクレオソームマップ解析から細胞間におけるヌクレオソームの形成位置の保存性と多様性が明らかになり、多くの遺伝子の転写開始点付近では位置取りが高度に一致しています。この結果はヌクレオソーム位置取りが転写調節に深くかかわっていることを強く示唆しています。それでは転写開始点付近におけるヌクレオソームの位置取りは異なる生物種においても共通性があるでしょうか?我々はこの疑問に応えるため、系統進化上広い範囲の菌類を対象として、ヌクレオソーム位置取りの比較研究を行っています。今回、担子菌類としては世界初のヌクレオソームマップをゼンマイ寄生菌類Mixia osmundaeを用いて作成し、子嚢菌類と担子菌類におけるヌクレオソーム形成様式の異同について比較しました。その結果、ヒストンに巻きついているDNA長には種特異性があり、系統進化上近縁であっても相違が見られることを示しました。また、ヌクレオソーム間のリンカーDNA領域における塩基配列の偏りを示し、ヌクレオソーム形成領域における塩基配列の偏りと対照性があることを明らかにしました。この偏りはヌクレオソームマップが作成された生物に共通性があり、DNA配列とヒストンタンパク質の相互作用が進化の過程において広く継承されてきたことを示しています。
発表内容
菌類は“下等菌類”(ツボカビ類および接合菌類)と“高等菌類”に分類されます。“高等菌類”は有性胞子が内生である子嚢菌類と外生である担子菌類に分けられます。有性胞子の形成が確認できない“高等菌類”(いわゆる“不完全菌類”)も系統進化上は子嚢菌類か担子菌類に分けられます。ゼンマイ寄生菌類Mixia osmundaeはその発見記載より80年余りの間子嚢菌類に分類されてきましたが、1995年にこれまでに類似報告のないユニークな担子菌類であることを明らかにしました(Nishida et al., Can. J. Bot. 73: S660-S666)。しかし、その詳細な系統学的位置は不明のままです。そこで、今回、M. osmudnaeのゲノム塩基配列、RNAマッピング、網羅的転写開始点の決定、およびヌクレオソームマッピングを並行して解析いたしました。
その結果、ドラフトゲノム塩基配列として283コンティグ(約13.4 Mbp)を決定し、6726のタンパク質コード領域を予測いたしました。これらの予測タンパク質の類似配列検索の結果、本菌類がプクシニア菌亜門に属することを明らかにしました。
ヌクレオソームの位置については一つのヌクレオソーム(単一ヌクレオソーム)由来のDNA断片より約3000万、連続する二つのヌクレオソーム(単二ヌクレオソーム)由来のDNA断片より約2700万の位置を決定しました。それらの長さの分布は単一ヌクレオソームが132と150塩基の二つのピークを持ち、単二ヌクレオソームが300塩基のピークを持つことが分かりました。この長さの分布は糸状子嚢菌類Aspergillus fumigatusに近い結果であり、子嚢菌酵母Saccharomyces cerevisiaeとは大きく異なりました。なお、A. fumigatusは単一ヌクレオソームが135と150塩基のピーク、単二ヌクレオソームが285塩基のピークを持っており、M. osmundaeの単二ヌクレオソーム長よりも15塩基短いことがわかり、ヌクレオソームリンカーDNAの長さがM. osmundaeはA. fumigatusよりも若干長いことを示しています。
転写開始点上流から下流におけるヌクレオソーム形成位置のプロファイルでは、転写開始点に最も近いヌクレオソームの位置の保存度は極めて高く、転写開始点より離れれば離れるほど保存度が低くなる傾向を顕著に示しました。この特徴はゲノムワイドなヌクレオソームマップが作成された他の生物と共通でした。
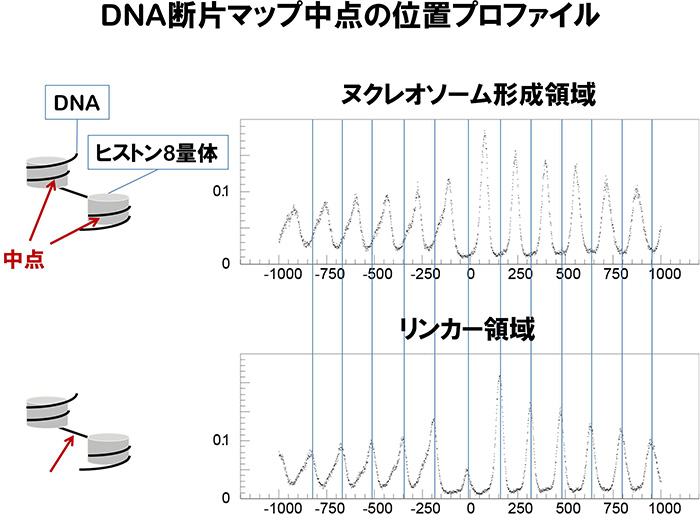
単一ヌクレオソームDNA断片の中心位置と単二ヌクレオソームDNA断片の中心位置のプロファイルを比較したところ、両者の振幅の様子が類似し、山と谷が一致していることが分かりました(図1)。すなわち、MNaseによるクロマチンの分解はほぼランダムに生じており、単二ヌクレオソームとして切られやすい位置が特定されているわけではないことを示しました。例外として、転写開始点から+1と-1の位置のヌクレオソーム由来のDNA断片は顕著に少なく、このDNA領域がMNaseにいち早く分解されたことを示します(図1)。
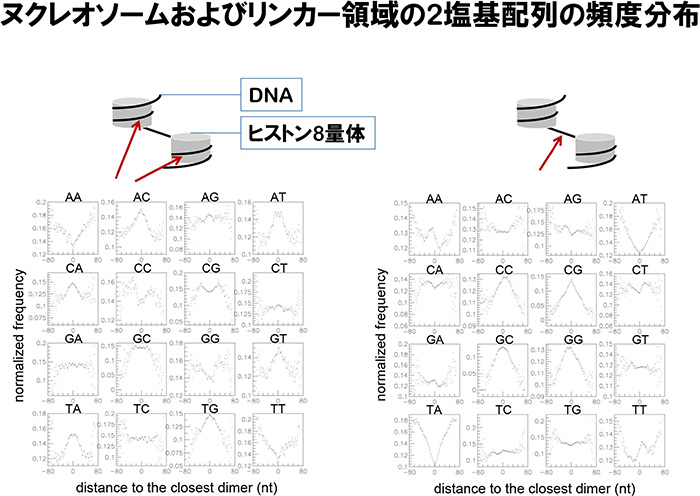
また、単一ヌクレオソームDNAの中心位置はヒストン8量体に結合している領域であり、単二ヌクレオソームDNAの中心位置はヌクレオソームリンカーDNAに位置していることから、それらのDNA配列の特徴を明らかにしました。リンカーDNA領域ではCC, CG, GC, GGの2塩基頻度が高く、AT, TAの頻度が低く、AT, TAの頻度が高いヌクレオソームDNA領域とは対照的であることを示しました(図2)。この特徴はヒトのヌクレオソーム研究で報告された結果とよく一致しており、真核細胞生物の進化において維持され、継承されてきたことを示しています。最近、遺伝子プロモータ領域におけるヌクレオソームの位置がATP依存的に配置されていることが明らかにされていますが、その後のヌクレオソーム形成においてはゲノム塩基配列の特徴に従い配置されていると考えられます。
発表雑誌
- 雑誌名
- Open Biology (Volume 2, Article ID 120043)
- 論文題名
- Characteristics of nucleosomes and linker DNA regions on the genome of the basidiomycete Mixia osmundae revealed by mono- and dinucleosome mapping
- 著者
- Hiromi Nishida, Shinji Kondo, Takashi Matsumoto, Yutaka Suzuki, Hirofumi Yoshikawa, Todd D. Taylor, and Junta Sugiyama
- DOI
- 10.1098/rsob.120043
問い合わせ先
東京大学大学院農学生命科学研究科 アグリバイオインフォマティクス教育研究ユニット
微生物インフォマティクス・フォーラム
特任准教授 西田洋巳
E-mail: hnishida@iu.a.u-tokyo.ac.jp