新しいトリプトファン代謝経路を放線菌から発見
- 発表者
-
尾崎 太郎 (現 東京大学大学院農学生命科学研究科 特任助教;当時 東京大学大学院農学生命科学研究科応用生命工学専攻博士課程3年、日本学術振興会特別研究員(DC1))
西山 真 (東京大学生物生産工学研究センター教授)
葛山 智久 (東京大学生物生産工学研究センター准教授)
発表概要
バクテリアの一種である放線菌から、新しいトリプトファン代謝経路を発見しました。また、この代謝経路によってニトリル基を持った新しいインドール誘導体が生産されることも明らかにしました。
発表内容
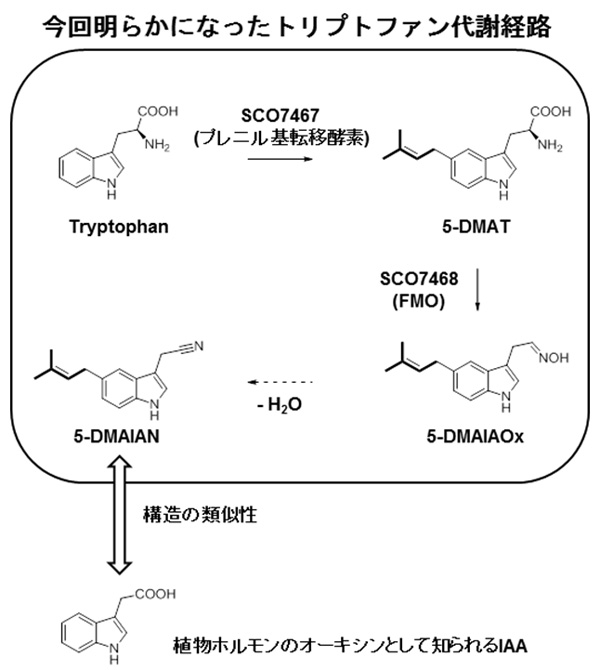
図 今回明らかにされたトリプトファン代謝経路。最終産物である5-DMAIANは植物ホルモンとして知られるIAAと類似した構造をしている。 (拡大画像↗)
放線菌は、ストレプトマイシンなどの抗生物質をはじめとする多様な二次代謝産物(注1)を生産する土壌細菌であり、産業上重要な微生物です。DNA配列の解読と解析技術の進歩によって、放線菌を含む様々な微生物ゲノムの配列情報が容易に入手できるようになり、一株の放線菌が数十もの二次代謝産物の生合成遺伝子を有することが明らかになりました。しかし、多くの生合成遺伝子が見出されるにもかかわらず、それぞれの放線菌からはわずか数種類の化合物しか同定されていないのが現状であり、多くの生合成遺伝子は発現していないか、または発現がきわめて弱いため、その機能がわかっていません。そのような機能未知の生合成遺伝子の中には、有用な生理活性を示す新規化合物の生合成に関与するものが多く存在すると期待され、近年その機能解析が盛んにおこなわれています。
今回、東京大学生物生産工学研究センターの葛山准教授の研究グループは、全ゲノム配列が公開されている放線菌Streptomyces coelicolor A3(2)に存在する、プレニル基転移酵素遺伝子SCO7467、およびフラビン依存型モノオキシゲナーゼ(FMO)遺伝子SCO7468を含む機能未知生合成遺伝子群に着目し、その機能を明らかにしました。これら二つの遺伝子をクローン化し、別の放線菌であるStreptomyces lividans TK23に導入することで、これらの遺伝子産物によって生合成される化合物は、新規構造を持つ5-ジメチルアリルインドール-3-アセトニトリル(5-DMAIAN)であることが明らかにされました(図)。
葛山准教授らは、トリプトファンを出発物質とする5-DMAIANの生合成経路の解明にも成功しました。その生合成経路では、初めに、プレニル基転移酵素であるSCO7467がトリプトファンにプレニル基を付加して5-ジメチルアリルトリプトファン(5-DMAT)を生成します。次に5-DMATは、SCO7468というFMOによって5-ジメチルアリルインドール3-アセタルドキシム(5-DMAIAOx)へと変換されます。類似の反応は植物由来のシトクロムP-450によって触媒されることがこれまでに報告されていましたが、FMOによって触媒される反応は初めての報告例です。5-DMAIAOxは最終的に脱水反応を受けることで5-DMAIANへと変換されると考えられます。
葛山准教授らは、今回明らかにした新たなトリプトファン代謝経路やその類似経路を担う遺伝子が多くの放線菌のゲノム上に存在することも見出しました。インドール骨格を持つ化合物は様々な生理活性を示すことが知られています。特に、インドール酢酸(IAA)は、オーキシン(注2)という植物ホルモンとしての生理活性を示します。今回発見した5-DMAIANもオーキシンに近い構造をしていることと、多くの放線菌が土壌細菌であることを考え合わせると、5-DMAIANのようなオーキシン類似化合物を介した放線菌と植物の応答現象の存在が想像されます。今後、5-DMAIANや類縁化合物の生理活性が明らかにされることで、新しい作物生産調節技術の開発につながるのではと期待されています。
本研究は、総合科学技術会議により制度設計された最先端・次世代研究開発支援プログラムにより、日本学術振興会を通して助成されたものです。
発表雑誌
- 雑誌名
- 「The Journal of Biological Chemistry 288: 9946-9956 (2013)」(4月5日号)
- 論文タイトル
- Novel tryptophan metabolism by a potential gene cluster that is widely distributed among actinomycetes.
- 著者
- Taro Ozaki, Makoto Nishiyama, and Tomohisa Kuzuyama
問い合わせ先
東京大学生物生産工学研究センター 細胞機能工学研究室
准教授 葛山智久
Tel: 03-5841-3073
Fax: 03-5841-8030
E-mail: utkuz@mail.ecc.u-tokyo.ac.jp
用語解説
- (注1) 二次代謝産物
- 生物の生育や生命活動の維持に直接は関与していない代謝産物の総称。
- (注2) オーキシン
- 植物の成長を促進する作用などを持つ植物ホルモンの総称。天然由来のオーキシンとしてインドール酢酸(IAA)がよく知られている。