シダ植物ではフェロモンとしても機能するジベレリン
- 発表者
-
中嶋 正敏(東京大学大学院農学生命科学研究科 応用生命化学専攻 准教授)
Park Seung-Hyun(東京大学大学院農学生命科学研究科 応用生命化学専攻 博士研究員)
発表概要
発芽や草丈をコントロールする植物ホルモン・ジベレリンは、シダ植物において別の個体の成長をコントロールする「フェロモン」としても機能していることが明らかとなりました。
発表内容
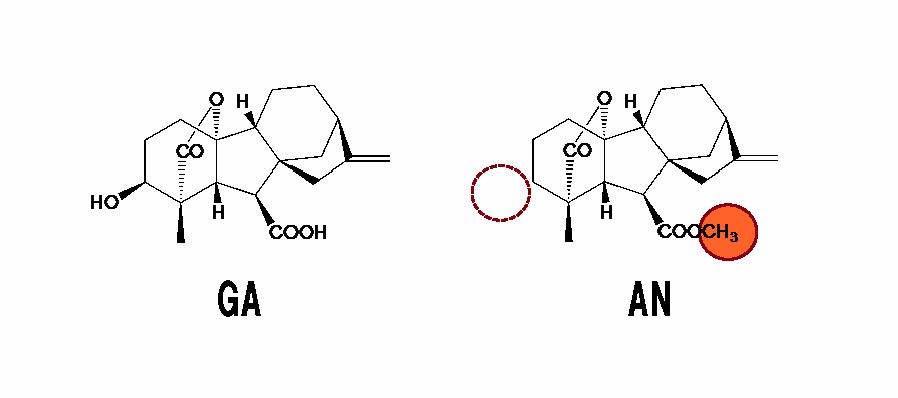
図1 代表的なジベレリン(GA)・造精器誘導物質(AN)の構造比較(拡大画像↗)
GAと比較してANは、① GAとして役立つために必須のOH基を持たない(波線部分)。これに加えて、② やはりGAとして役立つためには必須のカルボキシ(COOH)基に余分な「飾り(朱色部分)」がついて、構造が変化している。従って、ANはGAとしては役に立たない。
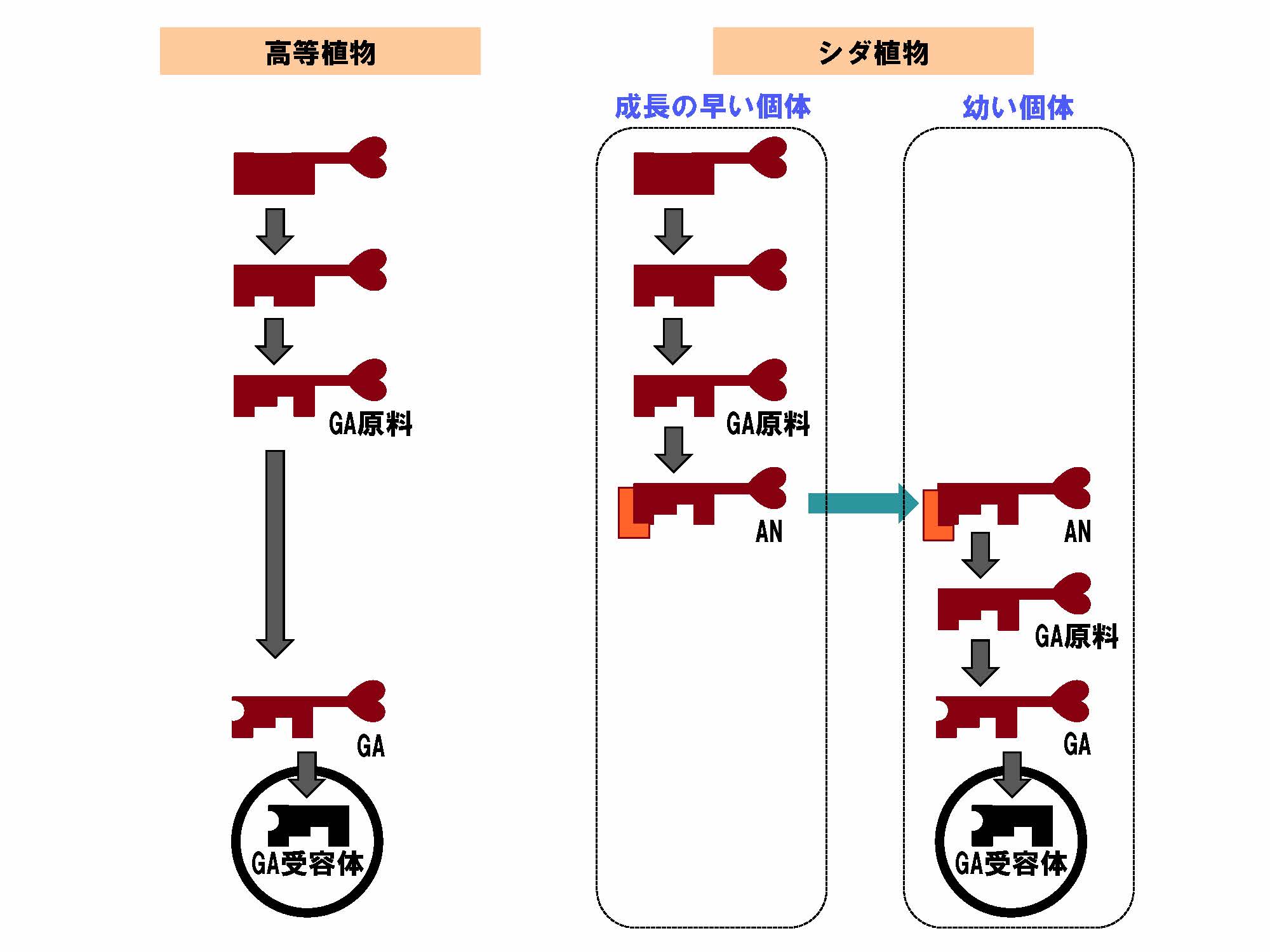
図2 シダ植物において分断が確認されたジベレリン(GA)の合成過程(拡大画像↗)
(左)一般的に高等植物で知られるGAの合成経路を、カギの形状変化になぞらえて模式的に表現した。すなわち、GA合成には複数の酵素が関与しているが、それら各酵素のはたらきにより段階的に構造が変化して、最終的にカギ穴(受容体)に合うカギ(GA)が作られる。ここでは、GAの直接の原料となる分子を「GA原料」と表記する。
(右)対してシダ植物では、集団の中で比較的成長の早い個体においてGA原料までの合成が盛んである。しかし、GAへの変換反応を司る酵素が存在しないため、GA合成はこの段階で中断される。その代わりに「飾り(朱色部分)」をつける反応を経て造精器誘導物質(AN)を合成する。ANは個体の外へ分泌され、幼い個体の表面に付着する。「飾り」がはずれる反応が生じてGA原料の構造に戻る。幼い個体ではGAへの変換反応を司る酵素が活発にはたらいているため、GAが合成される。
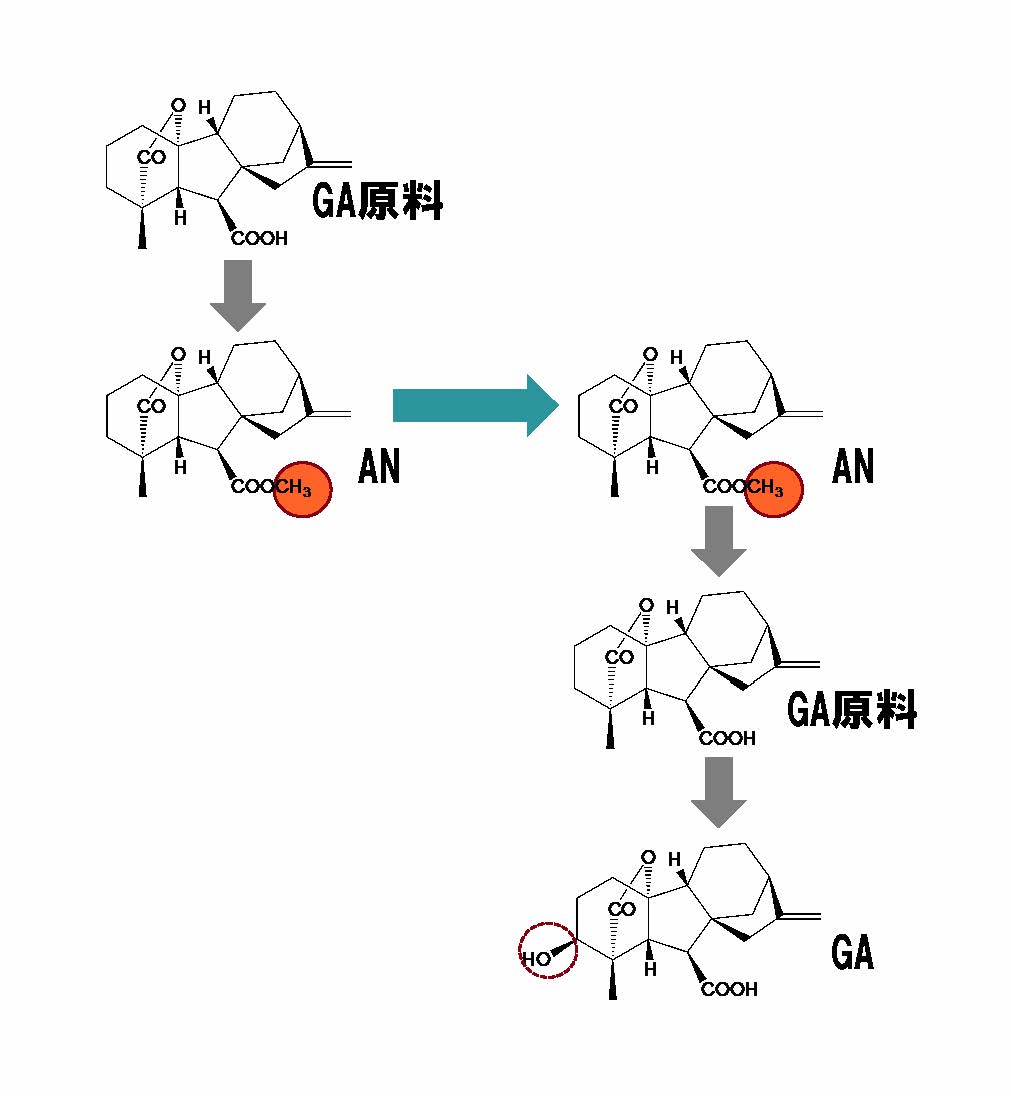
図3 GA合成過程の分断を橋渡しする造精器誘導物質(AN)と実際の構造変化(拡大画像↗)
比較的成長の早い個体において、GAの直接の原料となる分子(GA原料)が合成されたのち、カルボキシ基がメチル修飾(朱色部分)されて、ANとなる。幼い個体へ移動したANは修飾部分が脱離して、再びGA原料に戻る。GA合成の最終過程を司る酵素のはたらきにより水酸基(波線部分)が導入されてGAとなる。ANは、分断された2つの過程をつなぐ『橋渡し』役と表現できる。
ジベレリン(GA)は種子発芽や茎の伸長、花器官の発達など、植物自身の様々な成長をコントロールする植物ホルモン(注1)として知られ、例えばブドウやイチゴなどを処理対象の農産物として、生産の現場で広く利用されています。これまでの研究から、植物の体内でどのような過程を経てGAが合成されるか、また、GAからの信号はどのように伝わるか、それらの仕組みが概ね判明しています。他方、高等植物に比べて古くから地球上に存在していたと考えられるシダ植物では体内からGAが見つかりますが、未だにその役割は明確になっていません。一方、シダ植物には構造がGAにとてもよく似ている別の物質AN(アンセリディオーゲン、日本語では造精器誘導物質、図1)が存在して、比較的幼い個体(前葉体)におけるオスの器官(造精器と呼ばれています)の分化を誘導することが知られていました。
今回、カニクサと呼ばれるシダ植物を用いて、新たに以下の点が明らかとなりました。
①オス器官への分化誘導は、ANだけでなくGAでも生じるが、ANの方が誘
導活性が高い。
②GAの信号が伝わる仕組みをANも使う。ただし、ANはGAの受容体に直
接結合できない。
③GA受容体の機能を阻害したら、ANは誘導活性を示さなくなる。
④GA合成の最終段階を司る酵素の機能を阻害した場合も、ANは誘導活
性を示さなくなる。
⑤GA合成の最終段階を司る酵素は成長の早い個体では存在せず、GA合
成が中断される。
⑥水の中では、GAに比べてANはシダ植物の表面に効率良く付着する。
⑦水の中で幼い個体にANを与えると、GAの原料(⑤酵素の基質)が合成さ
れる。
⑧最終段階を司る酵素(⑤)は幼い個体のみで存在する →原料があれば
GAが合成される。
これらの情報を合わせると、シダ植物が持つ巧妙な仕組みが見えてきます(図2・3)。
[A] シダ群落中、成長の早い個体ではGA合成が中断され、(GAの原料とし
て利用されないように)飾りをつけることでANを合成する。ANは個体の
外へ放出される。
[B] 水を介して移動したANは、まだオス・メスの性が決まっていない周りの別
の幼い個体の表面に効率よく辿りつき、AN分子内の「飾り」部分をはず
してGAの原料に戻る。
[C] 幼い個体では、GA合成の最終段階を司る酵素がはたらいており、ANか
ら「飾り」部分がはずれることにより生じたGA原料を用いて、GAを合成
する。
[D] 結果、GAは受容体と結合して分化に必要な信号が伝わり、幼い個体に
おけるオス器官の分化が誘導されている、と説明することが可能となりま
した。
シダ植物においてGAが、他個体の成長を制御する「フェロモン(注2)」として機能していたことについては、高等植物でもGAが花器官の発達に重要なはたらきをしていることと考え合わせると、オス・メスの性に関わるコントロールが最も古くから備わっていたGA本来の機能であって、よく知られる種子発芽や茎の伸長に対するコントロールは、実は進化の過程で後天的に付け加わった機能と考えることができます。
発表雑誌
- 雑誌名
- 「Science」(24 October 2014, VOL. 346, Issue #6208)
- 論文タイトル
- Antheridiogen determines sex in ferns via a spatiotemporally split gibberellin synthesis pathway.
- 著者
- Junmu Tanaka, Kenji Yano, Koichiro Aya, Ko Hirano, Sayaka Takehara, Eriko Kouketu, Reynante Lacsamana Ordonio, Seung-Hyun Park, Masatoshi Nakajima, Miyako Ueguchi-Tanaka and Makoto Matsuoka
- DOI番号
- 10.1126/science.1259923
- 論文URL
- http://www.sciencemag.org/lookup/doi/10.1126/science.1259923
資金援助
本研究は、平成23年度文部科学省科学研究費補助金・新学術領域研究「ゲノム・遺伝子相関 ー新しい遺伝子分野の創成ー(領域代表 高山誠司)」、計画研究、遺伝子の重複・変異が生み出す新しい成長メカニズム(代表 松岡 信・分担 中嶋正敏)を始めとする公的研究資金のサポートを受けて遂行されました。
問い合わせ先
東京大学大学院農学生命科学研究科 応用生命化学専攻 生物制御化学研究室
准教授 中嶋 正敏
Tel:03-5841-5192
Fax:03-5841-8025
研究室URL:http://pgr.ch.a.u-tokyo.ac.jp
用語解説
- 注1 植物ホルモン
- 植物の体内で合成され、微量で自身の成長・分化を制御する低分子化合物の総称。
- 注2 フェロモン
- 生物の体内で合成され、微量で同種・他個体の成長・分化を制御する低分子化合物の総称。