コケにおける化学物質を利用した防御システムを発見 - 植物のストレス応答進化の解明につながる成果 -
- 発表者
-
岡田 憲典(東京大学生物生産工学研究センター 准教授)
川出 洋(東京農工大学大学院農学研究院 准教授)
宮本 皓司(帝京大学理工学部 研究員)
宮崎 翔(東京大学大学院農学生命科学研究科 特任研究員)
貝沼 遼介(東京農工大学大学院農学府 大学院生)
木村 穂乃香(東京農工大学大学院農学府 大学院生)
藤原 薫(東京大学生物生産工学研究センター 大学院生)
夏目 雅裕(東京農工大学大学院農学研究院 教授)
野尻 秀昭(東京大学生物生産工学研究センター 教授)
中嶋 正敏(東京大学大学院農学生命科学研究科 准教授)
山根 久和(帝京大学理工学部 教授)
波多野 友(岡山理科大学理学部 大学院生)
野崎 浩(岡山理科大学理学部 教授)
林 謙一郎(岡山理科大学理学部 教授)
発表のポイント
◆蘚類ハイゴケが生産するアレロパシー物質モミラクトンの生合成遺伝子を単離し、酵素活性および生物学的な機能を明らかにした。
◆下等植物の蘚類における低分子の化学物質を用いた誘導的なストレス耐性機構の存在を明らかにした初めての例である。
◆植物の化学物質を用いたストレス耐性機構のプロトタイプと考えられ、植物における生存戦略の進化的起源解明への貢献が期待される。
発表概要
東京大学生物生産工学研究センターの岡田憲典准教授と東京農工大学の川出洋准教授および岡山理科大学の林謙一郎教授らの研究グループは、東京大学大学院農学生命科学研究科、帝京大学理工学部と共同で、蘚類に分類されるコケ植物ハイゴケ(Hypnum plumaeforme)が生産するテルペノイド化合物モミラクトンの生合成の鍵となるHpDTC1遺伝子を単離し、その酵素機能とストレス応答性を明らかにしました。モミラクトンは、高等植物のイネにおいて抗菌性化合物のファイトアレキシンとして誘導的に生産されることが知られている生理活性物質です。これまでに、ハイゴケは周りの植物の生育を阻害する他感作用の原因物質としてモミラクトンを常に生産していることが報告されていましたが、今回、ハイゴケがカビなどの病原菌による感染を含む外界からのさまざまなストレスに応答してモミラクトン生合成遺伝子HpDTC1の発現を高め、誘導的にモミラクトンを増産していることがわかりました。このことは、高等植物がもつモミラクトンなどの低分子化学物質を武器とした病原菌に対する抵抗性発現システムのプロトタイプを、ハイゴケなどの下等コケ植物が持っていたことを意味します。植物の巧妙な生存戦略である化学物質を介した抵抗性発現システムの進化を解き明かす第一歩です。
発表内容
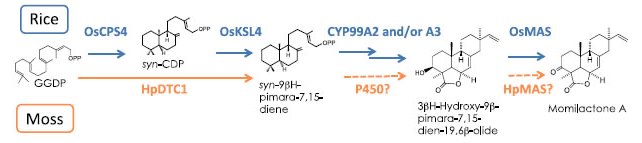
図1.ハイゴケとイネのモミラクトン生合成経路
ジテルペン系のファイトアレキシンであるモミラクトンは、GGDPという炭素数20のテルペン分子から、2段階の化学反応を経てピマラジエンに変換されたのちさらに数段階の酸化反応により生産される。上段のイネでは2つの酵素遺伝子によりGGDPからの2段階の環化反応が進むが、ハイゴケではHpDTC1の1つの酵素によって、2段階の反応が触媒される。(拡大画像↗)
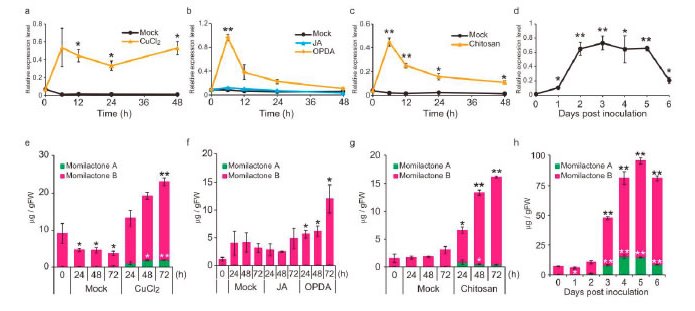
図2.さまざまなストレスにより誘導をうけるHpDTC1遺伝子
ハイゴケにおいてモミラクトン生産の鍵となる初発の環化反応をになうHpDTC1の遺伝子発現は、塩化銅やキトサンなどのエリシター処理だけでなく、病原菌であるB. cinerea菌の感染によっても誘導される。また、植物ホルモンのジャスモン酸では誘導を受けないが、その前駆体であるOPDAによる誘導を示す。HpDTC1の遺伝子発現誘導に合わせて、モミラクトンの生産も高まる。(拡大画像↗)
研究の背景
イネの抗菌性化合物として知られるジテルペン型ファイトアレキシン(注1)のモミラクトンは、蘚類ハイゴケにおいても他感作用(注2)を与える物質としてその生産が報告されていましたが、生合成遺伝子の情報は全くありませんでした。進化的にかけ離れたイネとハイゴケ(注3)において、同一化合物であるモミラクトンがどのように生合成されるのかは、低分子の化学物質を利用した植物の抵抗性システムを考える上でとても興味深い謎でした。
研究の経緯
モミラクトンはイネとハイゴケにおいて、周辺の植物に対する他感作用で働く成長抑制物質(アレロパシー物質)として生産されていることがわかっていました。一方で、イネにおいてはいもち病菌の成育を抑える抗菌性物質として、菌の感染時に誘導的に生産されることもこれまでの研究により明らかにされていました。本研究グループは、ハイゴケの生産するモミラクトンもカビなどの病原菌に対する抵抗性において機能しているのではないかと考え、まずハイゴケのモミラクトン生合成における鍵遺伝子を単離し、その機能を明らかにすると同時に、モミラクトン生産誘導における遺伝子発現の実態とその生物学的な意味の解明に取り組みました。
研究の内容・意義
ハイゴケにおけるモミラクトン生産は、塩化銅などの重金属ストレスで生産誘導を受けることが明らかにされていました。そこで、本研究グループは、塩化銅処理時に誘導を受ける遺伝子群をRNA-seq(注4)により探索し、その中からイネのモミラクトン生合成における鍵酵素であるコパリル二リン酸合成酵素とピマラジエン合成酵素の2つの酵素遺伝子が1つにまとまった構造をもつジテルペン環化酵素遺伝子HpDTC1を発見しました。大腸菌を用いた組換え酵素生産により試験管内反応で組換えHpDTC1がモミラクトン生合成の出発物質となる炭素数20のゲラニルゲラニル二リン酸を2段階の環化反応によってモミラクトンの前駆体となる炭化水素のピマラジエンにまで変換することを明らかにしました。
HpDTC1遺伝子は、塩化銅やキトサン処理の他、Botrytis cinerea(注5)などの病原菌の感染ストレスにより、mRNAの発現が劇的に高まることが分かりました。しかし、高等植物のイネにおいてモミラクトン生合成遺伝子の発現を誘導することが分かっている植物ホルモンのジャスモン酸(JA)には、HpDTC1の発現およびモミラクトン生産の誘導を引き起こす能力がないことが分かりました。さらにJAの前駆体物質であるオキソフィトジエン酸(OPDA)にはHpDTC1の発現とモミラクトンの生産の誘導効果が見られることも明らかにしました。ハイゴケを含むコケ類からはJAが検出されないことから、ハイゴケにおけるモミラクトンの生産誘導にはOPDAのシグナルが利用されている可能性を示しました。
HpDTC1のプロモーター領域をレポータ遺伝子であるGUSにつなぎ、そのプロモーターの発現をモデルコケ植物であるヒメツリガネゴケで観察したところ、ハイゴケにおけるHpDTC1の発現誘導と同様に、さまざまなストレスによってプロモーターの発現が誘導を受けることを明らかにしました。このことは、モミラクトン生産などのストレス応答が、蘚類のコケで保存された共通のシグナル伝達システムで駆動されている可能性を強く示唆するものです。
今後の予定・期待
本研究グループは、現在、ハイゴケにおけるHpDTC1遺伝子以外のモミラクトン生合成遺伝子の単離を進めており、ハイゴケのモミラクトン生合成経路の解明を目指しています。また、HpDTC1のプロモーターを材料として、ハイゴケにおけるモミラクトン生産誘導機構を転写レベルで明らかにする研究も進めて行きます。本研究で明らかにしたハイゴケの塩化銅誘導性遺伝子の中には、多くの転写制御因子が含まれており、その中には、モミラクトン生産に関わる因子も含まれている可能性があります。これらの転写制御因子の解析を進め、すでに明らかにされているイネのモミラクトン生産制御因子との比較を行うことで、植物の化学物質を介した抵抗性発現機構における転写制御の進化についても解き明かされることが期待されます。
発表雑誌
- 雑誌名
- 「Scientific Reports」
- 論文タイトル
- HpDTC1, a Stress-Inducible Bifunctional Diterpene Cyclase Involved in Momilactone Biosynthesis, Functions in Chemical Defence in the Moss Hypnum plumaeforme
- 著者
- Kazunori Okada, Hiroshi Kawaide, Koji Miyamoto, Sho Miyazaki, Ryosuke Kainuma, Honoka Kimura, Kaoru Fujiwara, Masahiro Natsume, Hideaki Nojiri1, Masatoshi Nakajima, Hisakazu Yamane, Yuki Hatano, Hiroshi Nozaki, Ken-ichiro Hayashi (Sci. Rep. 6, 25316;2016)
- DOI番号
- 10.1038/srep25316
- 論文URL
- http://www.nature.com/articles/srep25316
問い合わせ先
東京大学大学生物生産工学研究センター 環境保全工学研究室
准教授 岡田 憲典
Tel:03-5841-3070
Fax:03-5841-3070
研究室URL:http://park.itc.u-tokyo.ac.jp/biotec-res-ctr/kampo/index.html
用語解説
- 注1 ファイトアレキシン
- ファイトアレキシンは、植物がさまざまなストレスに応じて生産する化合物。植物種毎にさまざまな種類の化合物が存在する。イネのファイトアレキシンは化学構造や生合成経路の違いから、ジテルペン系のモミラクトン類、ファイトカサン類とフラボノイド系のサクラネチンに分類される。今回研究の対象としたファイトアレキシンはジテルペン系のモミラクトン。
- 注2 他感作用
- 植物が他の植物の生長を抑える物質(アレロケミカル)を放出したり、あるいは動物や微生物を退けたり引き寄せたりする効果の総称。英語ではAllelopathy(アレロパシー)といわれる。
- 注3 ハイゴケ
- ハイゴケ科ハイゴケ属のコケで蘚類に分類される。ほふく性で這うようにふえることから「這い苔」と命名された。アジアで幅広く見られる苔で、日本では園芸用に流通している。
- 注4 RNA-seq
- RNAの配列情報をDNAに変換し、そのDNAの配列を読むことにより、もとのRNAの配列を決定する手法。次世代シークエンサーによる高速シーケンスを用いてこの配列決定を行うことにより、遺伝子の発現状況を網羅的に解析する。今回の研究材料であるハイゴケのようなゲノム情報が不明な生物においても遺伝子発現情報を得ることができる。
- 注5 Botrytis cinerea
- 貴腐ワインにおいてブドウの糖度を高めるカビとして知られるが、幅広い植物に寄生する植物病原菌。果実や花の灰色かび病を初め、斑点病、立枯病などを引き起こす。