「イネの化学防御システムにおける遺伝子クラスターの進化軌跡を同定」
- 発表者
-
宮本 皓司(帝京大学理工学部 研究員)
藤田 雅丈(国立遺伝学研究所植物遺伝 研究員)
Matthew R. Shenton (国立遺伝学研究所植物遺伝 研究員)
明石 翔太(東京大学生物生産工学研究センター 実験研修生)
菅原 千都 (山形大学大学院農学研究科 大学院生)
坂井 亜莉里(山形大学大学院農学研究科 大学院生)
堀江 清孝(茨城大学大学院農学研究科 大学院生)
長谷川 守文(茨城大学大学院農学研究科 准教授)
川出 洋(東京農工大学大学院農学研究院 准教授)
三橋 渉(山形大学大学院農学研究科 教授)
野尻 秀昭(東京大学生物生産工学研究センター 教授)
山根 久和(帝京大学理工学部 教授)
倉田 のり(国立遺伝学研究所植物遺伝 教授)
岡田 憲典(東京大学生物生産工学研究センター 准教授)
豊増 知伸(山形大学大学院農学研究科 教授)
発表のポイント
◆野生イネゲノム情報と化学防御物質ファイトアレキシンの生産を調べることで、ファイトアレキシン生合成遺伝子クラスターの進化過程を明らかにした。
◆栽培種のイネOryza sativaで知られる化学防御物質の生合成遺伝子クラスターは、人による育種以前の野生イネにおいて既に構築が始まっていたことを示す進化論を提唱した。
◆イネのファイトアレキシン生合成遺伝子クラスターは、野生イネのゲノム上の特定の位置において構築されたと考えられ、ゲノム領域の制御メカニズム解明への進展が期待される。
発表概要
東京大学生物生産工学研究センターの岡田憲典准教授と山形大学の豊増知伸教授を中心とする研究グループは、国立遺伝学研究所、帝京大学理工学部と共同で、Oryza属のジテルペン抗菌性化合物であるファイトアレキシンの生産を担うゲノム上の遺伝子クラスター形成進化について、栽培化前の複数の野生イネを用いた解析により、その構築起源と考えられる進化過程を明らかにしました。ファイトアレキシンは、イネの栽培種であるO.sativaにおいて抗菌性化合物として誘導的に生産されることが知られている生理活性物質ですが、この生合成遺伝子群がゲノム上でクラスターを構成することを以前の研究で明らかにしておりました。今回、栽培イネが持つ化学防御物質の生産能と遺伝子クラスターの保持が、育種の過程で有用形質として選抜されたのではなく、イネが野生種(雑草)だった時代において進化的なゲノム構成の過程で獲得されていたことがわかりました。さらに、ファイトアレキシンの生産能と遺伝子のクラスター化の間には相関があり、不完全なクラスターを保持した野生イネ系統では、ジテルペン型ファイトアレキシンの生産が認められませんでした。興味深いことに、ファイトアレキシンを生産しない野生イネ系統のゲノム上のクラスター構成位置には、祖先遺伝子と考えられる一部のシトクロームP450水酸化酵素遺伝子が座乗しており、あたかもクラスター構築のための予定地であったかのようなゲノム構造であることも明らかにしました。このことは、現在の栽培イネが装備しているファイトアレキシンを武器とした病原菌に対する抵抗性発現システムは、野生イネのゲノム進化のなかで遺伝子クラスターとして獲得されてきたことを意味します。どうしてクラスター化する必要があり、イネにとってどのようなメリットがあるのか?今回明らかにした成果には、イネが長い年月をかけて構築した抵抗性発現システムにおける遺伝子のクラスター化の謎を考える上で重要なヒントが隠されていると考えています。
発表内容
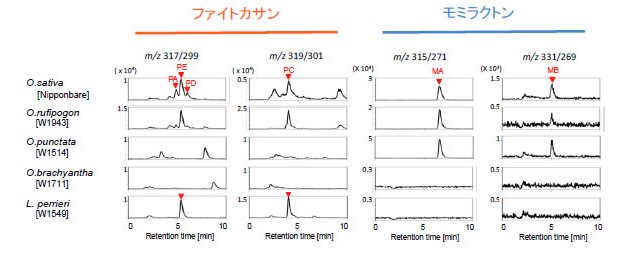
図1 野生イネとLeersia perrieriにおけるファイトアレキシンの生産性
ジテルペン型のファイトアレキシンであるモミラクトンとファイトカサンは、ゲラニルゲラニル二リン酸という炭素数20のテルペン分子から、2段階の化学反応を経てピマラジエンあるいはカサジエンに変換されたのちさらに数段階の酸化反応により生産される。O.punctataではファイトカサンの生産は認められないが、モミラクトンのみ生産されている。O.brachyanthaではいずれのファイトアレキシンも生産されない。Oryza属の近縁他属であるL.perrieriではファイトカサンのみが生産される。(拡大画像↗)
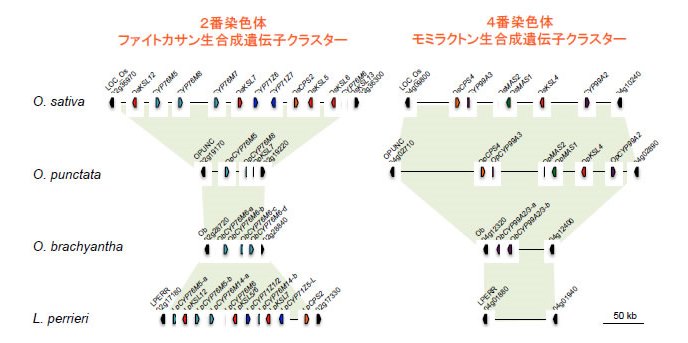
図2 野生イネとLeersia perrieriにみられるファイトアレキシン生合成遺伝子クラスター
ジテルペン型ファイトアレキシンの生産と生合成遺伝子クラスターの存在の有無は良く一致している。ファイトカサンの生産能を持たないO.punctataでは、2番染色体のクラスター領域に不完全な遺伝子配置が認められ、遺伝子クラスターの構築過程の様に見受けられる。ファイトアレキシン生産能を持たないO.brachyanthaにおいても、2番染色体と4番染色体にファイトアレキシン生合成遺伝子と相同性のある先祖遺伝子が存在している。Oryza属の近縁他属であるL.perrieriにおいて2番染色体にファイトカサン生合成遺伝子クラスターが存在することから、Oryza属で一旦ファイトカサン生産能を遺伝子クラスターの崩壊により失ったか、L.perrieriとは独立してファイトカサン生合成遺伝子クラスターを形成した2つの可能性が考えられる。(拡大画像↗)
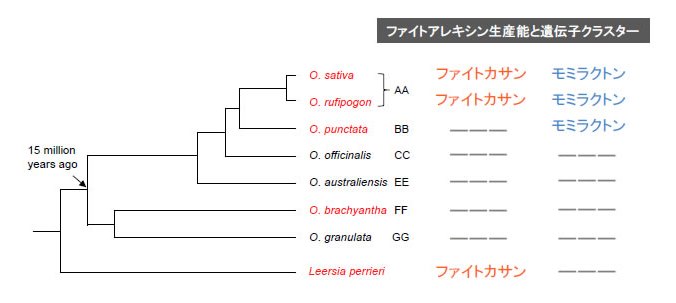
図3 野生イネとLeersia perrieriの進化系統樹とファイトアレキシン生産能の関係
二倍体のOryza(イネ)属には、細胞遺伝学的な形質、ゲノムDNA交雑法、遺伝子の配列などの違いによる6種類の ゲノムタイプ(AA、BB、CC、EE、FFおよびGG)が存在し、進化系統分類に用いられる。Oryza属におけるファイトカサンとモミラクトンの生産能および遺伝子クラスターの獲得はBBゲノムタイプのO.punctataを境に起こったと考えられる。一方で、L.perrieriのファイトカサン生合成遺伝子クラスターは、Oryza属の分岐よりも前に存在しており、Oryza属とは異なる独自の進化をたどった可能性がある。(拡大画像↗)
研究の背景
イネ栽培種であるO.sativaにおいて、抗菌性化合物であるジテルペン型ファイトアレキシン(注1)の生合成遺伝子群がゲノム上でクラスターを構成することが知られておりました。高等植物における二次代謝物質の生合成遺伝子クラスター(注2)の存在は、ゲノム配列情報が明らかになるにつれてその報告が増加してきていますが、植物がなぜこのような遺伝子クラスターを構成するのか、どのようなメリットがあるのかについてはよくわかっていません。この点を解き明かす重要な情報として、イネが進化上いつ頃ファイトアレキシン生合成遺伝子クラスターを獲得したのかについては、興味深い謎でした。
研究の経緯
ジテルペン型ファイトアレキシンのモミラクトンはその抗菌活性により、イネの重要病害であるいもち病菌に対する抵抗性の付与に重要な役割を果たしていることが示されています。イネがモミラクトンを生産することはイネの生存戦略として十分理解できますが、同時にこの有用形質は我々人間にとっても水稲栽培において利用価値が高いものです。本研究グループでは、ファイトアレキシン生産能とその遺伝的な要素となるゲノム上の遺伝子クラスター構成が、人間によるイネの育種の過程で構築されてきた可能性を考えて、人による育種を受けていないイネ野生種におけるファイトアレキシン生産能を化合物レベルで明らかにすると共に、ゲノム情報を利用した遺伝子クラスターの保持と祖先遺伝子からの詳細な進化モデルの提唱を試みました。
研究の内容・意義
Oryza(イネ)属ついては、栽培種であるO.sativaからゲノム構造の類似性を基にタイプ分けされた約20種の野生イネが存在し、国立遺伝学研究所のナショナルバイオリソースプロジェクトOryzabaseにおいて系統維持と実験材料としての分与が行われています。そこで、本研究グループは、遺伝学的に栽培イネから離れている多数の野生イネにおけるファイトアレキシン生産能をLC-MSMS分析(注3)により調査し、2種のジテルペン型ファイトアレキシンであるモミラクトンとファイトカサンの両方を生産する系統、片方のみ生産する系統、あるいはどちらの生産能も保持しない系統を明らかにしました。さらに、それらの野生イネのゲノム情報を詳細に調べることで、栽培イネのゲノム上に見いだされるファイトアレキシン生合成遺伝子クラスターが野生イネにも存在し、ファイトアレキシンの生産性とよく一致することを示しました。
野生イネの中でも栽培イネのO.sativaに一番近縁であり直接の祖先種であるO.rufipogonでは、ジテルペン型ファイトアレキシンの生産性と遺伝子クラスターの構成において、栽培イネとの間にほとんど相違が認められませんでした。一方で、さらに進化的距離が離れたO.punctataにおいてはファイトカサン生産能が認められず、モミラクトン生産のみが確認されました(図1)。ゲノム上のクラスター構成を見てみると、モミラクトン生合成遺伝子クラスターについては栽培イネと同じく4番染色体上の同様な位置に保存されていましたが、2番染色体上のファイトカサン生合成遺伝子クラスターが存在すべき位置には、一部の生合成遺伝子のみが認められ、不完全な遺伝子クラスターであることが明らかになりました。また、さらに遠縁のO.brachyanthaにおいてはジテルペン型ファイトアレキシンの生産は認められず、二つの遺伝子クラスター領域には生合成の後半を担うシトクロームP450水酸化酵素の祖先遺伝子と思われる遺伝子のみが座乗していました。この結果はO.brachyanthaからO.punctataにいたる系統進化の過程で、4番染色体のモミラクトン生合成遺伝子クラスターが構築された可能性を示しています。さらに、Oryza属に最も近縁な他属であるLeersia perrieri についても調べたところ、興味深いことにファイトカサン生合成能を保持しており、遺伝子クラスターについても遺伝子数と配置には違いが認められるものの、2番染色体の同様な染色体の位置に存在することを発見しました(図2)。この結果から、ファイトカサンはOryza属よりも以前から遺伝子クラスターと共に存在しており、ある種のOryza属では一旦その生産能をクラスターの崩壊とともに失い、運良くこのクラスターを維持できた系統がO.rufipogonを経て現代の栽培イネに継承された可能性を強く示唆するものです(図3)。
今後の予定・期待
本研究グループは、現在、クラスター構成とファイトアレキシン生産能の異なる複数の野生イネについて、RNA-seq(注4)を用いた発現解析を進めています。このような生合成遺伝子クラスターの構成が、その領域に含まれる遺伝子の発現に与える影響を調べるため、特に、不完全な遺伝子クラスター領域における遺伝子の発現変動に着目しつつ、ファイトアレキシン生合成遺伝子クラスターの制御メカニズムの解明を目指しています。これまでに栽培イネにおいて明らかにしてきた転写制御因子が、野生イネにおいても同様な機能をはたしているのかどうかを掘り下げていくことで、イネの生存戦略である化学防御物質を介した抵抗性発現機構における転写制御の進化についても理解が進むことが期待されます。
発表雑誌
- 雑誌名
- 「The Plant Journal」
- 論文タイトル
- Evolutionary trajectory of phytoalexin biosynthetic gene clusters in rice.
- 著者
- Koji Miyamoto, Masahiro Fujita, Matthew R. Shenton, Shota Akashi, Chizu Sugawara, Arisa Sakai, Kiyotaka Horie, Morifumi Hasegawa, Hiroshi Kawaide, Wataru Mitsuhashi, Hideaki Nojiri, Hisakazu Yamane, Nori Kurata, Kazunori Okada*, Tomonobu Toyomasu*. (* Corresponding authors)
- DOI番号
- 10.1111/tpj.13200
- 論文URL
- http://onlinelibrary.wiley.com/doi/10.1111/tpj.13200/
abstract;jsessionid=7D7593C3322D84A3965514E580AD3331.f01t01
問い合わせ先
東京大学大学生物生産工学研究センター 環境保全工学研究室
准教授 岡田憲典
Tel:03-5841-3070
Fax:03-5841-3070
研究室URL:http://park.itc.u-tokyo.ac.jp/biotec-res-ctr/kampo/
用語解説
- 注1 ジテルペン型ファイトアレキシン
- ファイトアレキシンは、植物がさまざまなストレスに応じて生産する化合物。植物種毎にさまざまな種類の化合物が存在する。イネのファイトアレキシンは化学構造や生合成経路の違いから、ジテルペン型のモミラクトン類、ファイトカサン類とフラボノイド型のサクラネチンに分類される。ジテルペン型のファイトアレキシンは全て共通の前駆体であるゲラニルゲラニル二リン酸を経て生合成され、炭素数20の化合物である。
- 注2 生合成遺伝子クラスター
- 二次代謝化合物の中にはその生合成遺伝子が染色体上でクラスターを形成することがある。特にバクテリアにおけるオペロン構造がよく知られているが、高等植物においても生合成遺伝子クラスターの存在が数多く示されている。イネにおいては、2種のジテルペン型ファイトアレキシンに対応した2つの生合成遺伝子クラスターが2番染色体と4番染色体に存在する。
- 注3 LC-MSMS分析
- 液体クロマトグラフ-タンデム型質量分析計(LC-MSMS)を用いた有機化合物の同定・定量方法であり、LC部でカラム担体との親和性の差によって成分ごとに分離し、ターゲット化合物の分子質量と、特定の質量のみをさらに解離・フラグメント化した質量を、2段階の質量分析計で検出する分析手法。調微量の測定が可能であり、今回の研究のような既知化合物の定量分析にも適している。
- 注4 RNA-seq
- RNAの配列情報をDNAに変換し、そのDNAの配列を読むことにより、もとのRNAの配列を決定する手法。次世代シークエンサーによる高速シーケンスを用いてこの配列決定を行うことにより、遺伝子の発現状況を網羅的に解析する。今回の研究材料であるハイゴケのようなゲノム情報が不明な生物においても遺伝子発現情報を得ることができる。