最小オープンリーディングフレーム「AUG-stop」を介したリボソームを舞台とした新たな遺伝子発現制御機構の発見
発表のポイント
◆ リボソームは植物の栄養素の一つであるホウ素の細胞内の濃度が高くなると、その機能の一部を停止し、それに伴ってホウ素輸送体の合成を抑制しmRNAを分解することを発見した。
◆ 今回発見した応答機構は動物でも機能する普遍的なもので、様々な栄養や細胞内環境に応じた多くの遺伝子発現制御に重要な可能性があり、新たな生物機能制御の方策につながる。
発表概要
<研究の背景> 植物が生育するには土壌からの栄養吸収が不可欠で、吸収を担う輸送体(注2)をそれぞれの栄養に対して持っています。ホウ素は植物に必須な無機栄養素ですが、高濃度のホウ素は毒性を示すため、必要量に応じて吸収量を調節する必要があります。これまでの研究で転写やタンパク質分解を介した調節機構の一端が知られていましたが、シロイヌナズナのホウ素吸収に重要なNIP5;1については調節機構が明らかになっていませんでした。
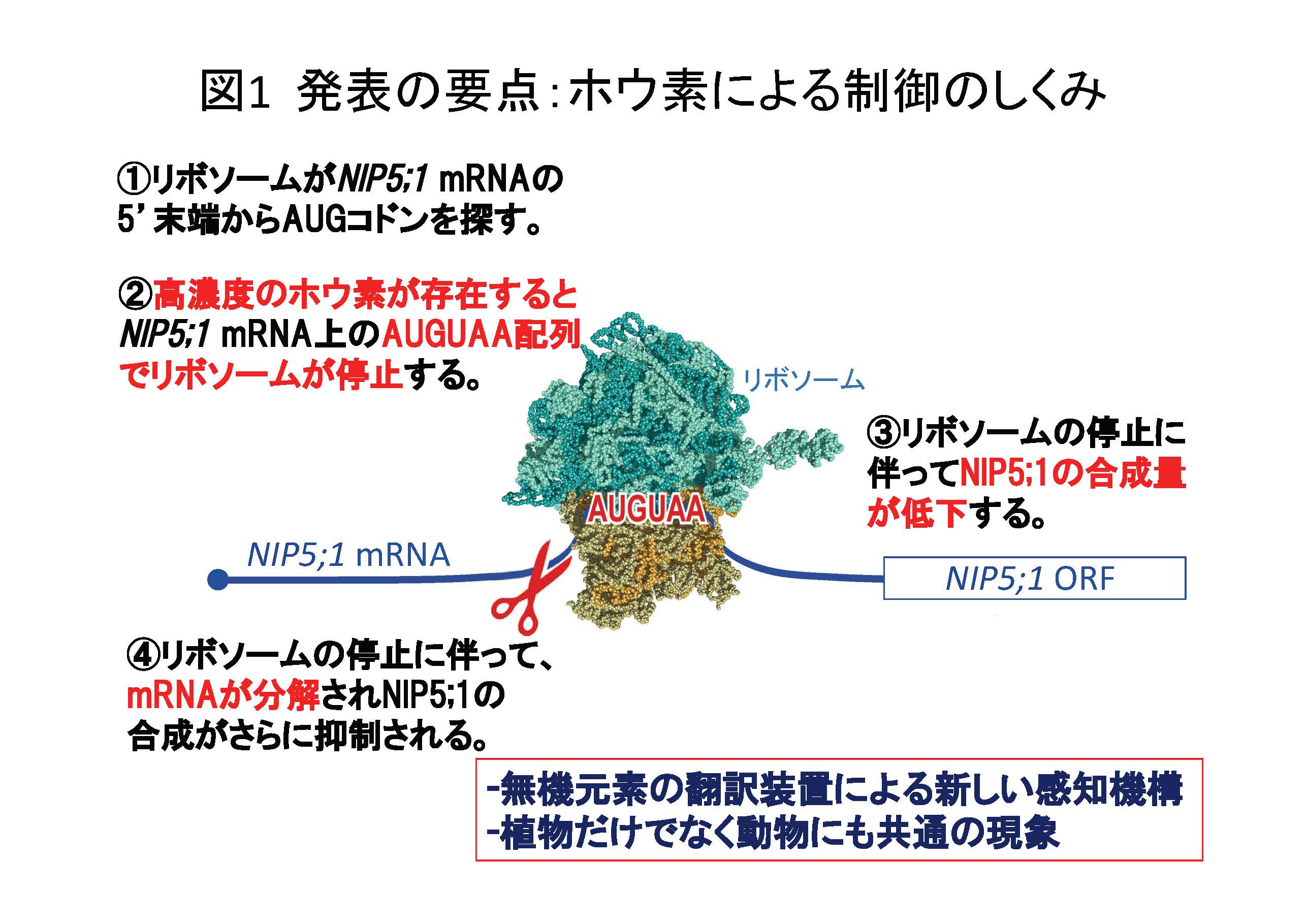
図1発表の要点:ホウ素による制御のしくみ(拡大画像↗)
<今回の成果> NIP5;1の栄養条件依存的な発現制御はこれまで全く注目されていなかった最も短いオープンリーディングフレーム上でのリボソーム停止を介した新たな仕組みで起こっていることが明らかになりました。(図1)
<研究手法・内容>
東京大学大学院農学生命科学研究科 藤原 徹教授北海道大学大学院農学研究院 内藤 哲教授らは、東京大学分子細胞生物学研究所、北海道大学理学研究院、北海道大学地球環境科学研究院、理化学研究所と共同で植物の生育に必要な栄養の吸収に関わるタンパク質の一つNIP5;1の栄養条件に応じた発現制御がこれまでに知られていないリボソーム機能を介したタンパク質合成阻害とmRNA分解によって起こることを明らかにしました。
より詳細には、細胞内のホウ素濃度が高いと、タンパク質合成装置であるリボソームがNIP5;1 mRNAを鋳型にタンパク質合成(翻訳)する過程でホウ素濃度が高いことを感知して一部の機能停止を起こすこと、この機能停止にはNIP5;1 mRNAに存在するAUGUAA(AUGは開始コドン(注3)、UAAは終止コドン(注4))という配列が必要であることを明らかにしました。このリボソームの機能停止に伴って、NIP5;1タンパク質合成が抑制されると共に、NIP5;1 mRNAの分解が起こり、NIP5;1の合成を素早く抑制することができると考えられます。また、このような制御はシロイヌナズナNIP5;1以外の遺伝子やイネやトウモロコシのNIP5;1に似た遺伝子でも起こること、 動物細胞などでも同様の制御が起こることを示しました。この制御機構が植物だけでなく、真核生物に普遍的に存在する可能性が考えられます。
また、この発見はホウ素輸送体NIP5;1の蓄積量の制御が細胞質のホウ素濃度を感知して起こっていることも示しています。栄養素がどのように感知されるかはほとんどわかっておらず、今回の成果は栄養の新たな感知機構を示すものです。
本研究はこれまでにない栄養の感知機構とタンパク質の蓄積制御機構を明らかにしたものです。このような機構は生物一般に存在し、また他の栄養素についても同様に栄養条件に応じてタンパク質の合成を制御している可能性が考えられ、これまでの生物の持つ環境や代謝に応じた遺伝子の発現制御機構の理解に新たな展開をもたらすものと考えています。
発表内容
<背 景>
ホウ素は植物の生育に必須な微量元素であり、土壌のホウ素が少ないと植物は欠乏症を発症します。一方で高濃度のホウ素は毒性があるため、ホウ素が過剰になっても生育障害を引き起こします。植物体内のホウ素濃度が適正に保たれることが農業生産や植物の生育に重要です。世界にはホウ素が少なすぎる土壌も、多すぎる土壌も分布しており、ホウ素欠乏症や過剰症を防止することは重要です。
nodulin-26 like protein (NIP) 5;1はシロイヌナズナのホウ酸輸送体であり、土壌からホウ酸を吸収するのに重要なタンパク質です。土壌のホウ素濃度が低いとNIP5;1 mRNAは多く蓄積し、多くのタンパク質が合成され、ホウ素の効率的な吸収を可能にします。この遺伝子を欠損させると、ホウ素欠乏条件では生育ができなくなります。また、NIP5;1のmRNAの蓄積はホウ素が十分に存在すると大幅に減少し、NIP5;1タンパク質が合成されないようにします。ホウ素が十分にある条件でホウ素を効率よく吸収するとホウ素毒性が顕在化する可能性があり、そうならない様に制御していると考えられます。
今回の研究では、この制御にタンパク質合成装置(リボソーム)が関与していることを明らかにしました。
<研究方法・成果>
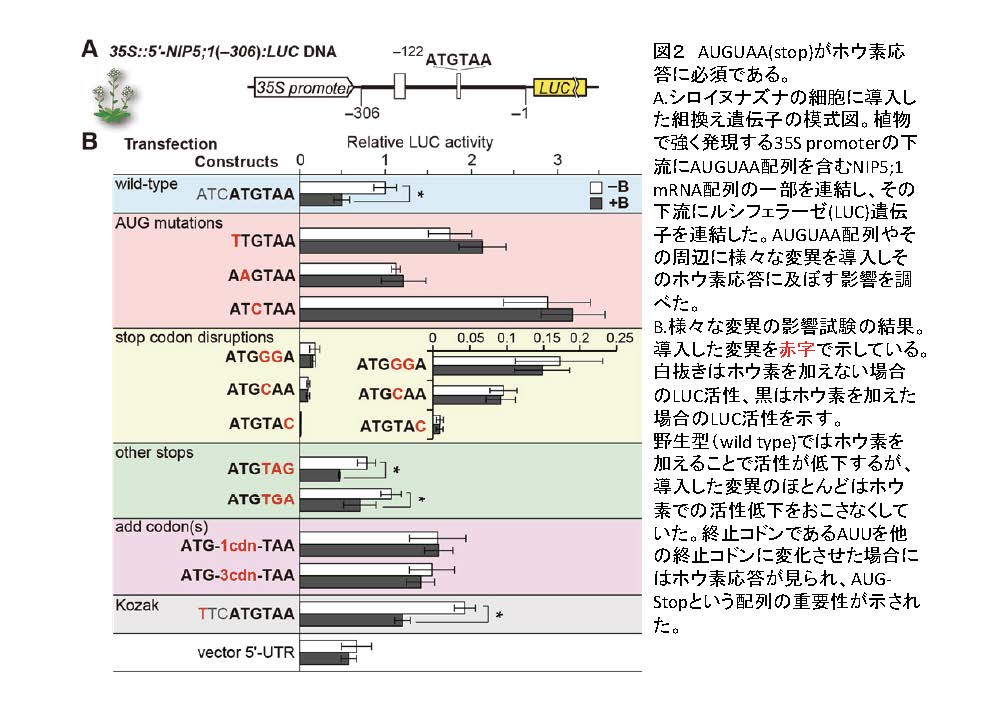
ホウ素条件に応じてNIP5;1のmRNAの蓄積がどのように制御されているかを明らかにするために、NIP5;1のmRNAのどの配列が制御に重要かを調べました(図2)。2011年の論文で既に私たちはNIP5;1のmRNAの蓄積の制御は、NIP5;1 mRNA上のNIP5;1をコードするオープンリーディングフレーム
(ORF)の上流側の領域を介して行われていることを明らかにしていました
(Tanaka et al. 2011)。本研究では、この領域の中のどの配列が制御に重要かを、配列を人為的に変化させたNIP5;1遺伝子を植物細胞に導入し、導入した遺伝子のホウ素による発現制御を調べることによって考察しました。その結果、AUGUAAという配列が不可欠であることが明らかになりました(図2)。AUGUAAという配列は開始コドン(AUG、注3)と終止コドン(UAA、注4)だけの組み合わせです。AUGを他のコドンに変化させるとホウ素に応じた発現制御が起こらなくなりました。終止コドンを終止コドン以外のコドンに変えても発現制御が起こらなくなりました。開始コドンと終止コドンの間にコドンを追加しても発現制御が起こらなくなりました。一方で、終止コドン(UAA)を他の終止コドン(UGAとUAG)に変えてもホウ素による発現制御が見られました。これらのことから、ホウ素による発現制御には開始コドンと終止コドンが連続していることが必要であることがわかりました。
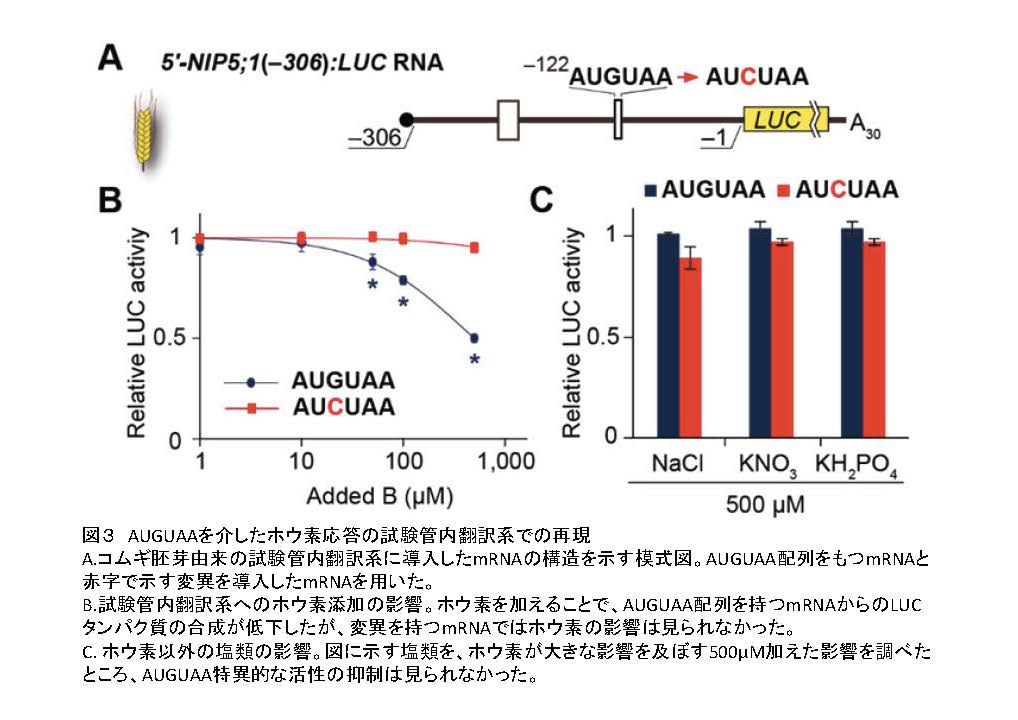
このような反応がどのように起こるかを知る手がかりとして、試験管内翻訳反応系でホウ素の効果を調べました(図3)。試験管内翻訳反応系は、リボソームなどのタンパク質合成装置をふくむ細胞抽出液で、反応液にmRNAを加えると、通常の細胞では細胞質で起こるリボソームによるタンパク質合成反応を再現できる実験系です。この実験系に、AUGUAA配列を持つmRNAを入れて翻訳反応をさせたところ、ホウ素を与えることで、翻訳反応が阻害されることが明らかになりました。このような阻害はAUGUAA配列を持たないmRNAでは起こりませんでした。また、ホウ素以外の塩類では反応は見られませんでした。これらの結果はホウ素に応じた制御が、細胞質で起こることを示唆しています。
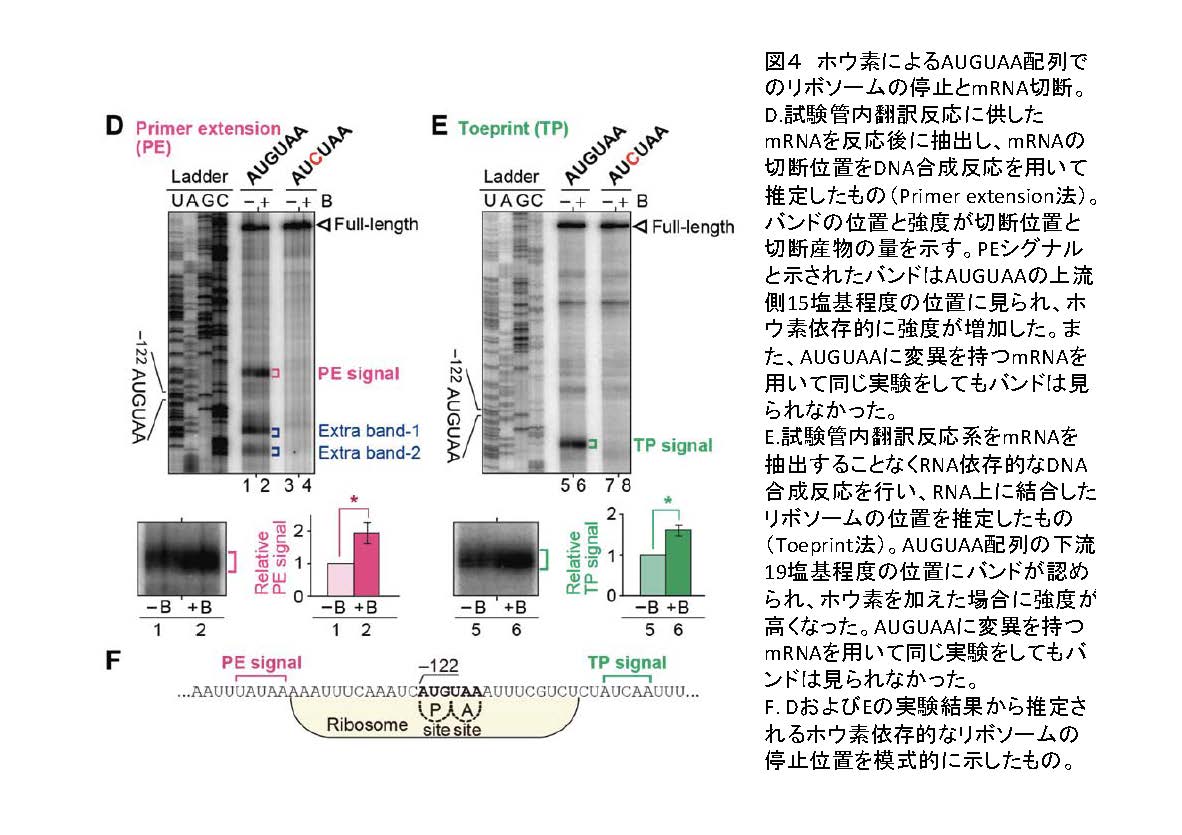
リボソームはmRNAの5’末端からAUGを探し、最初のAUGをみつけるとタンパク質の合成反応を始めると考えられています。AUGUAA配列に依存的なホウ素による制御にはリボソームが関与している可能性が考えられました。まずmRNAの切断点をプライマーエクズテンション法(注5)で解析しました(図4)。その結果、AUGUAAの上流15塩基付近で切断されていることが明らかになりました(図4)。さらに、リボソームの挙動を検討するために、トープリント試験を行いました。トープリント試験(注6)では、mRNA上でどの位置にリボソームが存在するかを推定することができます。ホウ素の濃度を変えてトープリント試験を行ったところ、ホウ素濃度が高いとAUGUAA配列でリボソームが停滞しやすくなることが明らかになりました(図4)。停滞しているリボソームは翻訳を行いません。タンパク質合成装置であるリボソームの挙動がホウ素により制御されており、このような制御がAUGUAA配列上で起こることが明らかになりました。
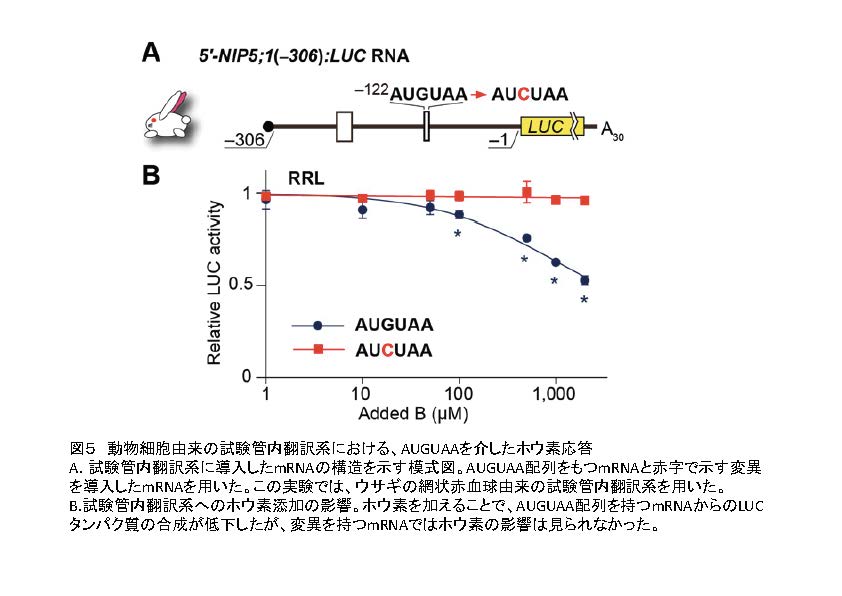
AUGUAAを介したホウ素による制御はシロイヌナズナのNIP5;1遺伝子以外の2つの遺伝子や、イネのNIP5;1相同遺伝子でも起こることを示しました。さらに、動物細胞由来の試験管内翻訳反応系で同様の応答が起こるかどうか調べたところ、ホウ素に対する応答が見られました(図5)。また、ヒト由来の細胞を用いてホウ素応答を調べたところ、ホウ素応答が見られました。これらのことはAUGUAA配列に依存したホウ素による発現制御機構は普遍的に存在することが明らかになりました。
遺伝子の中でタンパク質のアミノ酸配列を規定する領域をオープンリーディングフレーム(ORF)と呼びますが、AUGUAAはORFの中で考えうる限り最も小さいものです。この小さなORFがこのような機能を持つことを示した例はありません。本研究は、タンパク質合成装置が合成の過程で無機栄養を感知しその挙動を変えることを通じた遺伝子発現制御機構を初めて示したものです。このような制御は広く存在している可能性が考えられます。
発表雑誌
- 雑誌名
- 「The Plant Cell」on line版
- 論文タイトル
- “The Minimum Open Reading Frame, AUG-Stop, Induces Boron-Dependent Ribosome Stalling and mRNA Degradation”
- 著者
- Mayuki Tanaka, Naoyuki Sotta, Yusuke Yamazumi, Yui Yamashita, Kyoko Miwa, Katsunori Murota, Yukako Chiba, Masami Yokota Hirai, Tetsu Akiyama, Hitoshi Onouchi, Satoshi Naito*, and Toru Fujiwara*
- DOI番号
- 10.1105/tpc.16.00481
- 論文URL
- http://www.plantcell.org/content/early/2016/10/19/tpc.16.00481
問い合わせ先
<研究に関するお問い合わせ>
東京大学大学院農学生命科学研究科 応用生命化学専攻 植物栄養・肥料学研究室
教授 藤原 徹(ふじわら とおる)
Tel:03-5841-5104
Fax:03-5841-8032
E-mail:atorufu@mail.ecc.u-tokyo.ac.jp
用語解説
- 注1 オープンリーディングフレーム
- 遺伝子やmRNA上に存在する、タンパク質のアミノ酸配列を規定する領域。ORFと略記される。遺伝子にはタンパク質のアミノ酸配列を規定する領域以外に、mRNA合成を調節する領域などが存在しており、その中でタンパク質のアミノ酸配列を規定する領域のことを言う。
- 注2 輸送体
- 細胞膜等に埋まったタンパク質で特定の基質を膜の片側から反対側へ輸送させる能力を持つタンパク質の総称
- 注3 開始コドン
- オープンリーディングフレームの最初のコドンであり、翻訳を開始する位置を示す。メチオニンをコードするAUGが使われている。
- 注4 終止コドン
- オープンリーディングフレームの最後のコドンであり、翻訳を終了する位置を示す。UAA, UGA UAGの3種類があり、アミノ酸をコードしていない。
- 注5 プライマーエクズテンション法
- 文字通り、プライマーからDNA合成反応起こさせることによって鋳型となっているDNAやRNAの末端の位置を決める実験手法
- 注6 トープリント試験
- 試験管内翻訳系で反応させたmRNAをそのまま鋳型にしてプライマーからDNA合成反応起こさせることによってmRNAに結合しているリボソームやタンパク質の位置を調べる実験手法