ウイルスは植物の翻訳装置をハイジャックすることを発見
- 発表者
-
桂馬 拓也(東京大学 大学院農学生命科学研究科 生産・環境生物学専攻 博士課程3年、日本学術振興会特別研究員)
薦田 優香(酪農学園大学農食環境学群・循環農学類 講師/東京大学大学院農学生命科学研究科生産・環境生物学専攻
特任研究員:当時)
橋本 将典(ドイツ・マックスプランク植物育種研究所 客員研究員
/東京大学大学院農学生命科学研究科生産・環境生物学専攻日本学術振興会特別研究員)
煉谷裕太朗(東京大学大学院農学生命科学研究科 生産・環境生物学専攻 特任研究員)
鯉沼 宏章(東京大学大学院農学生命科学研究科 生産・環境生物学専攻 修士課程2年)
岩渕 望(東京大学大学院農学生命科学研究科 生産・環境生物学専攻 修士課程2年)
西田 萩子(東京大学大学院農学生命科学研究科 生産・環境生物学専攻 修士課程2年)
山次 康幸(東京大学大学院農学生命科学研究科 生産・環境生物学専攻 准教授)
難波 成任(東京大学 大学院農学生命科学研究科 生産・環境生物学専攻 教授)
発表のポイント
◆ウイルスが植物細胞の翻訳開始因子を乗っ取り利用することを明らかにしました。
◆ウイルスが翻訳開始因子を利用する分子メカニズムを解明しました。
◆翻訳開始因子を標的とした抗ウイルス薬開発やウイルス抵抗性品種の作出が期待されます。
発表概要
植物ウイルスは多様な植物に感染し萎縮や枯死などの症状を引き起こし、作物生産に甚大な被害を引き起こしますが効果的な化学薬剤は開発されておらず、防除が困難です。植物ウイルスはわずかな数の遺伝子しか持たないため、宿主の様々な遺伝子や分子装置を巧みに利用して宿主に感染します。ウイルスが利用する宿主の遺伝子を決定し、利用できないようにすれば、ウイルスが感染できなくなるため、宿主遺伝子の決定は重要です。
これまでの研究で一部の植物ウイルスが、植物の生存に必須な翻訳装置を構成する翻訳開始因子を利用することが分かっていました。そのうち翻訳開始因子4Eは遺伝子ファミリーを構成し、植物では3つのアイソフォーム(eIF4E、eIFiso4E、nCBP)を持ちます。いくつかの植物ウイルスがeIF4EとeIFiso4Eを利用することが分かっていました。しかし、植物ウイルスがnCBPを利用するかどうかは分かっていませんでした。
今回、東京大学農学生命科学研究科の桂馬拓也日本学術振興会特別研究員と難波成任教授らのグループは、ポテックスウイルス、ロラウイルス、カルラウイルスと呼ばれるグループのウイルスが植物に感染する際にnCBPを利用していることを明らかにし、その分子メカニズムを解明しました。これらの発見により植物ウイルスが翻訳開始因子4Eの全てのアイソフォームを利用することが示され、ウイルスが植物の翻訳装置を自身の都合がよいように利用している実態が明らかになりました。植物ウイルスが幅広く利用する植物遺伝子の解明により、翻訳開始因子を標的とする抗ウイルス薬の開発マーカー育種やゲノム編集技術を利用したウイルス抵抗性作物の作出が期待されます。
発表内容
ウイルスが植物に感染する際に、植物が持つ様々な遺伝子や分子装置を利用することが知られています。それらウイルスの感染に必須な植物遺伝子が変異している植物には、ウイルスは感染することができません。したがって、ウイルスの感染に必須な植物遺伝子を特定することにより、植物ウイルスに対する抵抗性を持つ作物の作出が可能になります。そのため、ウイルスが利用する植物遺伝子の決定は重要な意味を持ちます。
これまでの研究によって一部の植物ウイルスが翻訳開始因子4Eを利用することが明らかにされています。翻訳開始因子4EはDNAに刻まれる遺伝子情報をもとにタンパク質を合成する翻訳(注1)と呼ばれる反応において重要な役割をする因子で、遺伝子ファミリーを構成し、植物ではeIF4E、eIFiso4E、nCBPの3つのアイソフォーム(注2)を持つことが分かっています。このうちeIF4EもしくはeIFiso4Eが変異した植物にポティウイルス、ククモウイルス、カルモウイルスと呼ばれるグループの植物ウイルスが感染できないことから、これらのウイルスが、eIF4EとeIFiso4Eを利用すると考えられてきました。しかし、植物ウイルスが残るeIF4EアイソフォームであるnCBPを利用するかどうかは明らかでありませんでした。
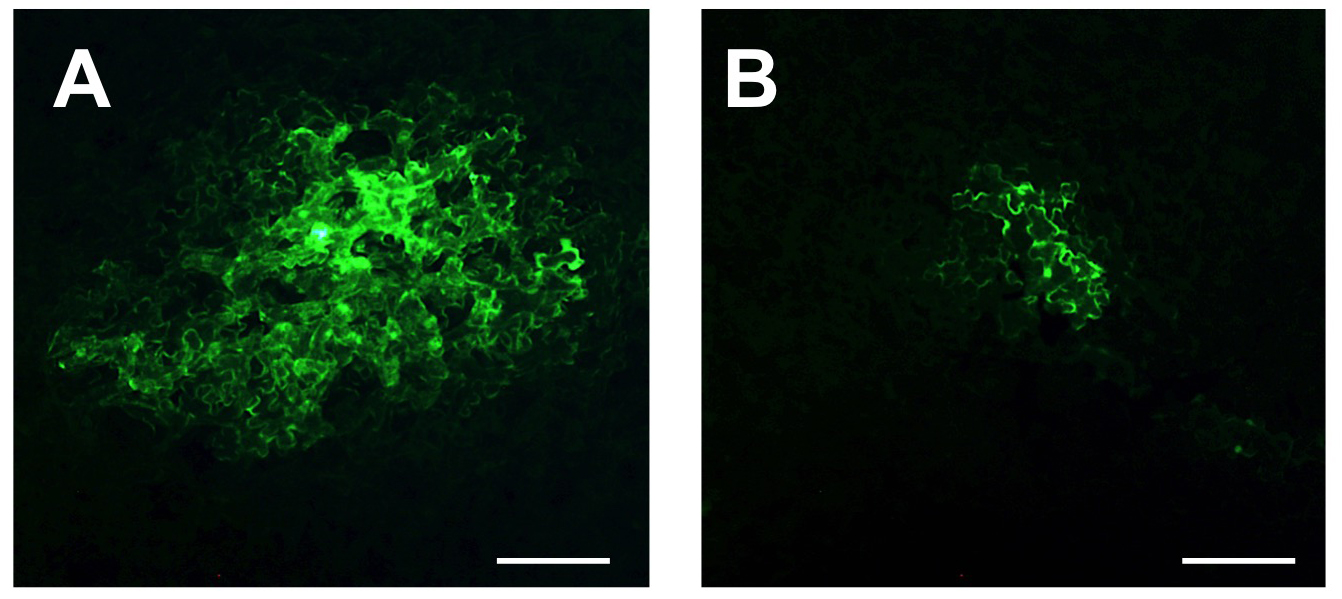
図1 nCBP変異体におけるウイルス細胞間移行の阻害
通常の植物(A)とnCBP遺伝子を変異させ機能を欠失させた植物(B)に光るタンパク質遺伝子(GFP)を導入したウイルスを接種して、その細胞間移行を観察した。通常の植物(A) に比べて、nCBP変異植物ではウイルスの細胞間移行が阻害されている。スケールバーは20 μm。(拡大画像↗)
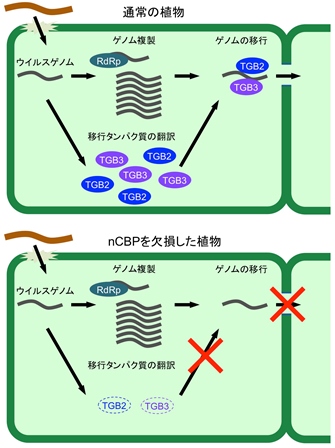
図2 nCBP変異体におけるウイルス細胞間移行の阻害メカニズム
通常の植物 (上) とnCBP遺伝子を変異させ機能を欠失させた植物 (下) におけるウイルスの感染の様子を模式的に示した。nCBPを変異させた植物では、移行タンパク質の蓄積量が減少し、ウイルスの細胞間移行が阻害されるが、植物の生育には影響がない。(拡大画像↗)
eIF4E、eIFiso4E、nCBP遺伝子がそれぞれ変異したシロイヌナズナ変異体にポテックスウイルスと呼ばれるウイルスグループのplantago asiatica mosaic virus (PlAMV)を接種したところ、nCBP変異植物では、PlAMVの感染が顕著に阻害されました。さらにnCBP変異植物では、ロラウイルスグループのlolium latent virusとカルラウイルスグループのpotato virus M(PVM)の感染も阻害されました。
次いで、nCBPがウイルスの感染にどのように関わるかを明らかにするために詳細な解析を行いました。nCBP変異植物では、PlAMVの初期感染細胞における複製(注3)は阻害されませんでしたが、感染細胞から隣接細胞への細胞間移行が阻害されていることが分かりました(図1)。さらに、nCBP変異植物では、PlAMVの持つ5つのウイルスタンパク質のうち、細胞間移行に必要な移行タンパク質triple-gene-block protein (TGB) 2およびTGB3の蓄積量が顕著に減少していることが分かりました。そのため、nCBP変異植物では、TGB2およびTGB3の蓄積量が減少したことでPlAMVの細胞間移行が阻害され、植物体への感染が阻害されたと考えられました(図2)。
③社会的意義・今後の予定など本研究により植物ウイルスが翻訳装置を構成する翻訳開始因子4Eの全てのアイソフォームを利用していることが明らかになり、その分子メカニズムが解明されました。このことはウイルスが植物の生存に必要不可欠な翻訳装置を巧みに利用し、感染を成立させていることを示します。今後ウイルスが翻訳装置やそこに含まれる因子をどのように利用するかを詳細に解析することで、ウイルスの感染戦略の理解につながるとともに、新たな抗ウイルス薬の標的発見の可能性が高まります。
また、本研究によりnCBP遺伝子が変異した植物ではポテックスウイルス、ロラウイルス、カルラウイルスの感染が阻害されることが明らかになりましたが、これらには農業上大きな被害をもたらす植物ウイルスが含まれます。ジャガイモ生産に甚大な被害を引き起こすpotato virus X、PVMや、世界中のトマトに甚大な被害をもたらし日本への侵入が警戒されているpepino mosaic virus、それぞれキク、ラン、ユリに重篤な被害を及ぼすchrysanthemum virus B、cymbidium mosaic virus、PlAMVなどが含まれます。しかし、これらの国内外で問題となっている植物ウイルスに対する効果的な防除法は確立されていません。nCBP遺伝子を標的としたマーカー育種やゲノム編集技術(注4)などにより、ウイルス抵抗性植物品種の開発につながることが期待されます。
なお、この研究は日本学術振興会科学研究費補助金、農林水産省農林水産業・食品産業科学技術研究推進事業の支援を受けて行われました。
発表雑誌
- 雑誌名
- 「Scientific Reports」(オンライン版:1月6日午後7時)
- 論文タイトル
- Deficiency of the eIF4E isoform nCBP limits the cell-to-cell movement of a plant virus encoding triple-gene-block proteins in Arabidopsis thaliana.
- 著者
- Takuya Keima, Yuka Hagiwara-Komoda, Masayoshi Hashimoto, Yutaro Neriya, Hiroaki Koinuma, Nozomu Iwabuchi, Shuko Nishida,Yasuyuki Yamaji and Shigetou Namba
- DOI番号
- 10.1038/srep39678
- 論文URL
- http://www.nature.com/articles/srep39678
問い合わせ先
東京大学大学院農学生命科学研究科 生産・環境生物学専攻 植物病理学研究室
教授 難波成任(なんば しげとう)
Tel:03-5841-5053
Fax:03-5841-5054
E-mail::anamba@mail.ecc.u-tokyo.ac.jp
研究室HP::http://park.itc.u-tokyo.ac.jp/ae-b/planpath/index.html
用語解説
- 注1 翻訳
- DNAに記録される遺伝情報は、まず「転写」とよばれる反応によりメッセンジャーRNA(リボ核酸)が合成され、その後に「翻訳」とよばれる反応によりタンパク質が合成されることにより、さまざまな生理反応が引き起こされます。「翻訳」はリボソームと呼ばれる巨大なタンパク質複合体を含む一連の翻訳装置によって行われますが、この翻訳装置には数多くの翻訳因子と呼ばれるタンパク質が含まれます。中でも、メッセンジャーRNAを引き寄せて翻訳を開始する起点となるのが翻訳開始因子4Eであり、翻訳において重要な役割を果たします。
ゲノム編集とは、DNA分解酵素を用いて遺伝子を自在に書き換える次世代技術です。DNA分解酵素としては、ジンクフィンガーヌクレアーゼ、TALEN、CRISPR/Cas9等が知られています。ゲノム編集を用いることで標的遺伝子を確実かつ迅速に改変できるため、交配と選抜を繰り返す上述の育種にかかる時間を劇的に短縮することが可能となります。ゲノム編集の際には、DNA分解酵素などの外来遺伝子を植物ゲノムに組み込む必要がありますが、改変後に非組み換え個体と交配することで、外来遺伝子を除去した個体が作出できます。すなわち、ゲノム編集により、遺伝子組換えの痕跡を持たない品種を迅速に作出することが可能となります。