統合失調症研究に新たな視点 -マウス成熟個体において認知機能を回復させることに成功-
- 発表者(※共同研究グループ)
-
糸原 重美 (理化学研究所 脳科学総合研究センター 行動遺伝学技術開発チーム チームリーダー)
吉田 崇将 (理化学研究所 脳科学総合研究センター 行動遺伝学技術開発チーム 研究員)
細谷 俊彦 (理化学研究所 脳科学総合研究センター 局所神経回路研究チーム チームリーダー )
中川 直 (理化学研究所 脳科学総合研究センター 局所神経回路研究チーム 研究員)
道川 貴章 (理化学研究所 脳科学総合研究センター 細胞機能探索技術開発チーム 研究員)
田中 三佳 (理化学研究所 脳科学総合研究センター 神経グリア回路研究チーム 研究員)
Thomas John McHugh (理化学研究所 脳科学総合研究センター 神経回路・行動生理学研究チーム チームリーダー)
Arthur Jyh-Yen Huang (理化学研究所 脳科学総合研究センター 神経回路・行動生理学研究チーム テクニカルスタッフ)
林 悠 (筑波大学 国際統合睡眠医科学研究機構(WPI-IIIS) 准教授)
(理研脳科学総合研究センター 行動遺伝学技術開発チーム 客員研究員)
柏木 光昭 (筑波大学大学院 人間総合科学研究科フロンティア医科学専攻 大学院学生)
桑原 正貴 (東京大学 大学院農学生命科学研究科獣医学専攻 教授)
安田 光佑 (東京大学 大学院農学生命科学研究科獣医学専攻 大学院生)
(理研脳科学総合研究センター行動遺伝学技術開発チーム 客員研究員)
要旨
統合失調症は幻聴・妄想などの「陽性症状」、意欲の低下・感情の平板化などの「陰性症状」、記憶力・注意力・情報処理能力などの機能が低下する「認知機能障害」を主な症状とする精神疾患です。従来の治療薬は、陽性症状の治療には有効ですが、陰性症状と認知機能障害に対しては十分な治療効果が得られず、患者の社会復帰を妨げる要因となっています。また発症には、「NMDA型グルタミン酸受容体(NMDA受容体)[2]」の機能低下が関わっていることが、古くから提唱されてきましたが、そのメカニズムは不明でした。
今回、共同研究グループは、脳の「視床髄板内核(ILN)[3]」におけるNMDA受容体の機能が生まれつき低下している遺伝子改変マウス(ILN変異マウス)を作製しました。そして、ILN変異マウスが記憶力・注意力・情報処理能力などの認知機能に障害を示し、活動量過多という陽性症状の一部に類似した症状を示すことが分かりました。また、統合失調症患者にみられる睡眠覚醒の障害と神経オシレーション活動[4]の異常を示すことも分かりました。これらの結果から、NMDA受容体の機能低下を引き金としたILN領域の機能異常が、統合失調症の病態に深く関わることが明らかになり、ILN変異マウスを統合失調症モデルマウスとして確立しました。さらにモデルマウスの成熟個体への遺伝子治療によりNMDA受容体の機能を正常に戻すことで、認知機能を“可逆的に”回復させることに成功しました。これまで、精神疾患の多くは脳の発達期に生じた不可逆的な要因が関与すると考えられていましたが、NMDA受容体の機能を後天的に回復させることが統合失調症の治療改善につながる可能性を示しました。
今後、モデルマウスを用いてILNの役割を明らかにすることにより、統合失調症の発症機序をより深く理解し、ILNのNMDA受容体を標的とした薬や遺伝子治療その他の手法による治療法の探索に役立つ知見が得られると期待できます。
本研究は、英国の科学雑誌『Translational Psychiatry』(2月28日付け:日本時間3月1日)に掲載されました。
背景
統合失調症は、およそ100人に1人の割合で発症する身近な精神疾患です。主な症状には、幻聴や妄想などの「陽性症状」、意欲の低下・感情の平板化などの「陰性症状」、記憶力・注意力・情報処理能力などの機能が低下する「認知機能障害」があります。従来の治療薬は、陽性症状の治療には有効ですが、陰性症状と認知機能障害に対しては十分な治療効果が得られず、患者の社会復帰を妨げる要因となっています。そのため、新たな治療法の探索に向けて、統合失調症のモデル動物の開発が求められていました。
神経伝達物質グルタミン酸の受容体の一種である「NMDA型グルタミン酸受容体(NMDA受容体)」の阻害薬は、健常者においても統合失調症に類似した症状を引き起こします。そのため、NMDA受容体に作用するグルタミン酸神経系の機能低下が統合失調症の病態に関与する可能性が古くから提唱されてきました。しかし、症状が脳のどの領域の機能低下によって引き起こされるのか、そのメ
カニズムは不明でした。
近年、統合失調症患者を対象とした機能的核磁気共鳴画像法(fMRI)[5]や死後脳の解析から、脳の深部にある視床の機能異常や、注意・覚醒を担う脳領域の一部と考えられる「視床髄板内核(ILN)」でNMDA受容体の発現量が減少していることが報告されています。
そこで共同研究グループは、“ILNにおけるNMDA受容体の機能低下によって、統合失調症の症状が引き起こされる”という仮説を立て、マウスを用いた遺伝学的手法により明確な因果関係を示すことを目指しました。
発表内容
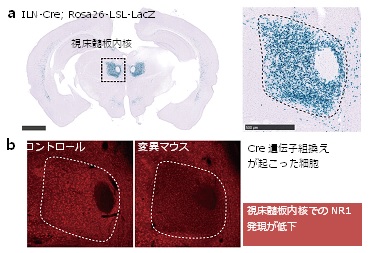
図1 新しい統合失調症モデルマウス(ILN変異マウス)の開発 a:マウスの脳切片におけるDNA組み換え酵素Creの空間的な発現パターン。発現部位を視覚的に確認できるレポーター系統(Rosa26-LSL-LacZ)を用いて、今回作製した視床随板内核(ILN)-Creマウスでは、Cre発現細胞(青色)がILNに限局していることを示している。 b:免疫組織化学法により可視化したNMDA受容体の必須サブユニットNR1の発現パターン。ILN-Creマウスと、NR1遺伝子のfloxアリルを持つマウスとの交配により生まれたILN変異マウス(ILN-Cre;NR1 flox/flox)では、コントロールマウス(NR1flox/flox)と比べて、ILNにおけるNR1の発現が低下していた。 (拡大画像↗)
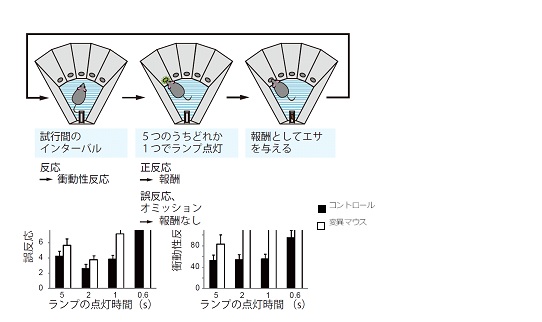
図2 新しい統合失調症モデルマウス(LIN変異マウス)で観察された注意・衝動性の障害
上: 5選択反応時間課題において、5つのランプのうち点灯した1つの穴に短時間で鼻先を入れ反応することで、報酬としてエサが獲得できることをマウスは学習する。
下: ランプの点灯時間を5秒、2秒、1秒、0.6秒とだんだん短くして難易度を高めると、ILN変異マウスでは、誤反応(ランプの点灯していない穴に反応する回数)や衝動性反応(ランプが点灯する前の待ち時間に衝動的に反応する回数)が増加したことから、注意力が低下しと衝動性が亢進していることが示された。
(拡大画像↗)
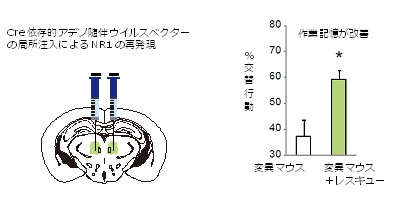
図3 遺伝子治療により新しい統合失調症モデルマウスの作業記憶が改善
左: アデノ随伴ウイルスベクターによる遺伝子治療。DNA組換え酵素のCre依存的にMNDA受容体の必須サブユニットであるNR1を発現するアデノ随伴ウイルスベクターを、統合失調症モデルマウスの視床髄板内核(ILN)に局所注入し、ILNにNR1遺伝子を再発現させ、NMDA受容体を回復させた。
右: Y字迷路試験で、統合失調症モデルマウスは交互に新しいアームを選ぶこと(交替行動)ができず、作業記憶(短期記憶)の障害を示したのに対して、遺伝子治療を行ったモデルマウスでは、交替行動がみられ作業記憶が改善した。
(拡大画像↗)
1)統合失調症の陽性症状の一部と認知機能障害を示すモデルマウスを開発
共同研究グループはまず、脳の視床髄板内核(ILN)におけるNMDA受容体の機能低下が統合失調症の病態に関わる可能性を検証するため、ILN領域で選択的にDNA組換え酵素Cre[6]を発現するトランスジェニックマウス[7] (ILN-Cre)系統を新たに開発しました(図1a)。このマウスをNMDA受容体の必須サブユニットであるNR1の遺伝子のfloxアリルを持つマウスと交配させて、ILNにおいてのみNMDA受容体を欠損する条件的変異マウス(ILN変異マウス、ILN-Cre; NR1 flox/flox)を作製しました(図1b)。
また、健常な被験者に突然強い聴覚刺激を与えると驚愕反応を示しますが、強い聴覚刺激の前に弱い聴覚刺激(プレパルス)を与えると、驚愕反応が大幅に抑制される現象(プレパルス抑制)が起こります。統合失調症患者ではプレパルス抑制の障害が報告されており、脳内の情報処理能力の障害を反映していると考えられています。げっ歯類の統合失調症の測定指標として行われる「プレパルス抑制試験」を行ったところ、ILN変異マウスはプレパルス抑制が弱いことが分かりました。
これら四つの結果から、ILN変異マウスは、記憶力・注意力・情報処理能力などの認知機能に障害を示すことが明らかになりました。
一方で、統合失調症患者は陽性症状の中で幻覚や妄想のほかに活動量過多を示します。げっ歯類では、陽性症状を検査するための指標として「歩行活動量」が用いられます。巣箱の中での歩行活動量を2日間にわたって測定したところ、ILN変異マウスは、夜間の歩行活動量が健常マウスよりも増加したことが分かりました。マウスは夜行性のため、ヒトと異なり昼間よりも夜間の活動が活発です。この結果から、ILN変異マウスは陽性症状の一部に類似した症状を示すことが明らかになりました。
このようにして、今回作製したILN変異マウスは、統合失調症の認知機能障害と陽性症状の一部を反映した新たな「統合失調症モデルマウス」として確立されました。
2)モデルマウスは睡眠覚醒の障害と神経オシレーション活動の異常を示す
睡眠には、レム睡眠[10](浅い眠り)とノンレム睡眠[10](深い眠り)があります。ヒトは入眠直後にノンレム睡眠に入り、次にレム睡眠へと移行します。眠りはこれら性質の異なる2種類の睡眠で構成され、約90分周期で一晩に4~5回、一定のリズムで繰り返されるといわれています。
統合失調症患者では、健常者に比べてノンレム睡眠が減少するという睡眠覚醒の障害が知られています。マウスは活動が活発な夜間でもときどき眠る習性がありますが、統合失調症モデルマウス(ILN変異マウス)では脳波の解析から、夜間にノンレム睡眠が減少し、覚醒している時間が増加することが分かりました。また、統合失調症患者では健常者と逆で、入眠後比較的すぐにレム睡眠に入るという特徴があります。同様のことが、モデルマウスでも昼間にみられました。これらの結果から、モデルマウスは夜間(ヒトでは昼間)に過覚醒状態にあり、昼間(ヒトでは夜間)にも統合失調症に類似した睡眠覚醒の障害を示すことが分かりました。
また、統合失調症患者では、大脳皮質における神経オシレーション活動の異常が報告されています。神経オシレーション活動は、注意力・記憶力・情報処理能力などの認知機能と関連することから、統合失調症での認知機能障害の発症機序に関わると考えられています。
脳波の分析から、モデルマウスでは夜間・昼間の両方で、レム睡眠・覚醒時にデルタ波(0.5-3Hz)、シータ波(4-8Hz)、アルファ波(8-12Hz)、ベータ波(13-30Hz)、ガンマ波(30-80Hz)の周波数帯での神経オシレーション活動が健常マウスよりも減弱しており、またノンレム睡眠時にガンマ波での神経オシレーション活動が健常マウスよりも減弱していることが分かりました。
以上の結果から、視床髄板内核(ILN)におけるNMDA受容体の機能低下が、統合失調症に類似した睡眠覚醒の障害や神経オシレーション活動の異常を引き起こすことが明らかになりました。
3)ウイルスベクター[11]を用いた遺伝子治療でモデルマウスの認知機能が回復
これまで多くの精神疾患の原因は、脳の発達期に生じた脳内の変異にあると考えられてきました。新たに作製した統合失調症モデルマウス(ILN変異マウス)でも、視床髄板内核(ILN)のNMDA受容体の機能低下により脳の発達が妨げられている可能性があります。そこで、ウイルスベクターを用いた遺伝子治療を行い、モデルマウス成熟個体においてILNにNMDA受容体を補うことで認知機能が回復されるか検証しました。
モデルマウス成熟個体に対して、ILNにおいてのみNMDA受容体を発現させることのできる特殊なアデノ随伴ウイルスベクター[11]を用いた遺伝子治療を行いました(図3左)。遺伝子治療後のモデルマウスをY字迷路試験により評価したところ、“可逆的に”作業記憶(短期記憶)を回復させることに成功したことが分かりました(図3右)。また、同じモデルマウスで、夜間の過活動性が抑制されたことも分かりました。
以上の結果は、統合失調症モデルマウスが示す行動異常の一部、つまり短期記憶力や陽性症状の一部が成熟個体において治療できることを示しており、ILNのNMDA受容体を標的とした治療、あるいはILNの活動を調整する手段が統合失調症に有効である可能性を示しています。
今後の期待
本研究では、視床髄板内核(ILN)におけるNMDA受容体の機能低下という機序に基づいた新たな統合失調症モデルマウスを樹立しました。またILNは、睡眠覚醒や神経オシレーション活動において、重要な役割を果たしていることが明らかになりました。このことは、統合失調症における睡眠覚醒や神経オシレーション活動の障害が、ILNの機能低下が原因となっている可能性を示しています。 今後、このモデルマウスを用いて、認知機能、睡眠覚醒、神経オシレーション活動におけるILNの役割を明らかにすることで、統合失調症の発症機序をより深く理解し、ILNのNMDA受容体を標的とした薬や遺伝子治療、さらに脳に直接的に刺激を与えるなどの治療法の探索に役立つ知見が得られると期待できます。