植物は2つの異なる応答機構によって低温ストレスに対する耐性を獲得している
- 発表者
-
城所 聡(東京大学大学院農学生命科学研究科 応用生命化学専攻 助教)
米田 考志(東京大学大学院農学生命科学研究科 応用生命化学専攻 博士課程2年;当時)
高崎 寛則(理化学研究所 環境資源科学研究センター 研究員;当時)
高橋 史憲(理化学研究所 環境資源科学研究センター 研究員)
篠崎 一雄(理化学研究所 環境資源科学研究センター センター長)
篠崎 和子(東京大学大学院農学生命科学研究科 応用生命化学専攻 教授)
発表のポイント
◆異常気象時や夜間などに起こる急激な温度低下に応答して植物の低温耐性を向上させる仕組みで働く転写因子(注1)を同定しました。
◆植物は2つの低温応答機構を用いて、季節変化などに起こる緩やかな温度低下と異常気象時や夜間などに起こる急激な温度低下を適切に感知し、耐性を獲得していることを明らかにしました。
◆寒波などによる急激な気温低下によって起こる作物の減収を防ぐための技術として応用されることが期待されます。
発表概要
東京大学と理化学研究所の共同研究グループは、植物が持つ低温ストレスに応答する2つの異なる仕組みを分子レベルで明らかにしました。植物は、低温、乾燥、高塩濃度といった環境ストレスを受けると、数多くの遺伝子の発現を変化させることにより耐性を獲得する機構を持っています。低温にさらされた植物は、DREB1と呼ばれる転写因子をコードする遺伝子の発現を強く誘導することで、耐性獲得に働く遺伝子群を強く働かせ低温耐性を獲得します。DREB1遺伝子(以下、イタリックで表記された単語は遺伝子を示す)の発現誘導に関わる因子は複数報告されていますが、それらがどのような仕組みで働いているのかは不明のままでした。今回、篠崎和子教授らの共同研究グループは、植物は季節変化などに起こる緩やかな温度低下と異常気象時や夜間などに起こる急激な温度低下とを、別々の機構で感知して耐性機構を働かせていることを明らかにしました。異常気象時や夜間の急激な温度低下時にはCAMTAと名付けられた転写因子が機能しており、季節変化などに起こる緩やかな温度低下では、植物の時計関連遺伝子が働いていることを示しました。前者は昼夜に関わらず機能しますが、後者は昼間のみに働いていることも明らかにしました。この研究により、植物の低温ストレスに対する感知機構の理解が進むとともに、寒波などによる急激な温度低下に対する耐性を向上させた作物の開発への応用が期待されます。
発表内容
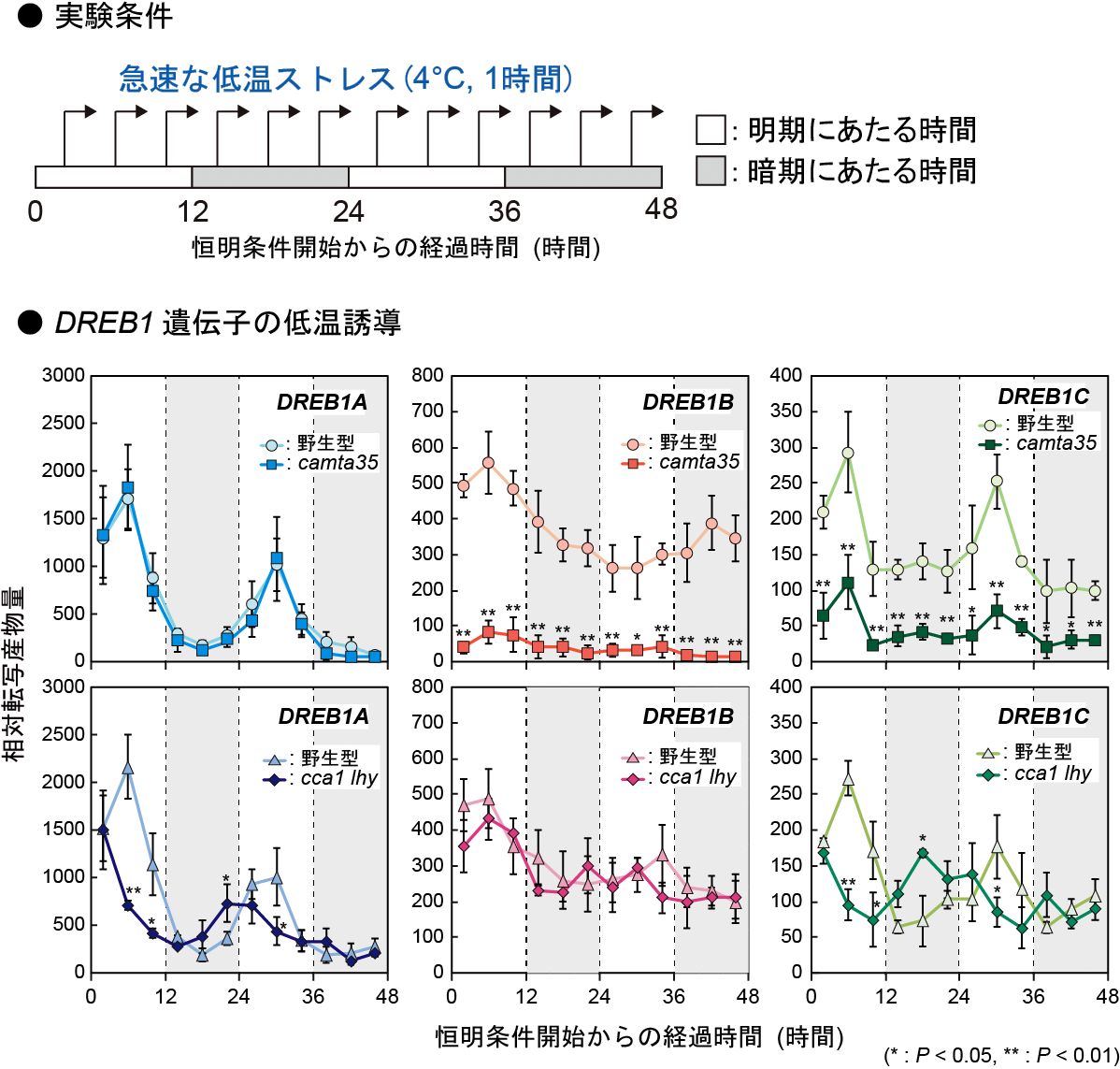
【図1】 DREB1AとDREB1C遺伝子の低温誘導は概日リズムを示し、昼間にのみ強く誘導されるのに対して、DREB1Bは昼間にも夜間にも強く誘導されます。CAMTA3とCAMTA5の変異体(camta35)ではDREB1BとDREB1Cの低温誘導が低下します。また、概日時計の中心的な構成因子である転写因子CCA1とLHYの変異体(cca1 lhy)では、DREB1AとDREB1Cの発現誘導に変化が見られたのに対して、DREB1Bでは変化が見られませんでした。(拡大画像↗)
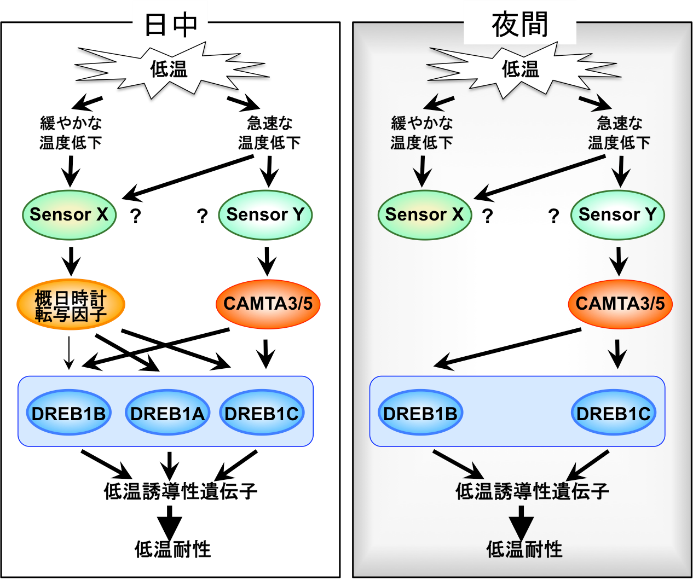
【図2】 昼間にはCAMTA3、CAMTA5と概日時計の転写因子の両方が機能することで温度低下の速さに関わらずDREB1の発現が誘導されます。それに対して、夜間には概日時計の転写因子が発現しておらずCAMTAだけが機能するため、急速に温度が低下するとDREB1の発現が誘導されます。(拡大画像↗)
地球レベルの環境劣化が進み、異常気象による農業被害が深刻化しています。さらに、地球温暖化の影響によって、急激な温度変化や干ばつなどが頻発するようになると予測されています。このため、環境ストレスに高い耐性を持つ作物の育種が重要となっています。植物は、環境ストレスを受けると、さまざまな遺伝子の発現を変化させることにより耐性を獲得します。DREB1遺伝子は、植物の低温ストレスに応答した耐性の獲得機構において中心的な役割を持つ転写因子をコードしています。DREB1転写因子は、植物が環境ストレス耐性を獲得するために機能する100以上の遺伝子の働きを活性化させることができることから、DREB1を高発現させた植物体は低温や凍結を含むさまざまな環境ストレスに対して高い耐性能を獲得します。DREB1遺伝子の発現は、植物が低温にさらされるとすぐに誘導されることから、DREB1遺伝子の低温誘導は、植物が低温に応答して耐性を獲得するための最初のステップであり、その機構の解明は植物の低温ストレスの感知機構の解明につながると推測されます。また、得られた結果は低温ストレス耐性作物の作出にも役立つと期待されます。モデル植物のシロイヌナズナにはDREB1A、DREB1B、DREB1Cの3つのDREB1遺伝子があり、これらの遺伝子の発現誘導に関わる因子は複数報告されていましたが、それらがどのような仕組みでDREB1遺伝子の発現を制御しているのかは不明のままでした。
今回、本共同研究グループは、シロイヌナズナのDREB1C遺伝子のプロモーター中の低温誘導に関わる配列に結合する転写因子であるCAMTAを単離して、その機能に着目して解析をおこないました。多重変異体を用いた解析から、6つのシロイヌナズナのCAMTAのうち、CAMTA3とCAMTA5が低温ストレス下でのDREB1BとDREB1Cの発現誘導を制御していることがわかりました。DREB1BとDREB1Cの発現は、急激な温度低下が起こった時にはCAMTA3とCAMTA5によって強く誘導されます。その一方で、緩やかな温度低下が起こった場合にはCAMTAの有無に関わらず発現が誘導されませんでした。このことから、CAMTA3とCAMTA5は急激な温度低下に反応して機能することが示されました。一方で、DREB1AはCAMTAによる制御を受けておらず、温度低下の速さに関わらず強く誘導されました。
このCAMTAによる制御機構の役割を調べるために、DREB1の低温誘導性に関わると考えられているもう1つの因子である概日時計(注2)による制御との関係性を調べました。DREB1の低温誘導性は概日時計によって制御されており、日中には強く誘導されるのに対して、夜間にはあまり誘導されないと報告されていました。しかし我々の解析の結果、DREB1AとDREB1Cの低温誘導は概日リズム(注3)を示し、日中に強く誘導されるのに対して、DREB1Bは昼夜問わずに強く誘導されることが明らかになりました。また、概日時計の中心的な構成因子である転写因子CCA1とLHYの変異体では、DREB1AとDREB1Cの発現誘導に変化が見られたのに対して、DREB1Bでは変化が見られませんでした(図1)。これらの結果から、DREB1AとDREB1Cの低温誘導は概日時計に制御されるが、DREB1Bの低温誘導は概日時計の制御をほとんど受けないことがわかりました。3つのDREB1遺伝子の低温時の発現誘導は、それぞれ異なる制御を受けており、DREB1Aは概日時計による制御、DREB1BはCAMTAによる制御、DREB1Cは概日時計とCAMTAの両方による制御を受けていることが明らかになりました(図2)。
以上の結果より、植物のDREB1を介した低温応答には、温度低下速度によって2つの異なる経路が働いていることが明らかになりました。1つは、急激な温度低下時にCAMTAによって活性化される経路で、この制御は昼夜問わずにDREB1の発現を誘導します。もう1つはCCA1やLHYといった概日時計関連の転写因子によって制御される経路で、こちらはCCA1やLHYが発現している日中に温度低下速度に関係なく活性化されますが夜間は働きを示しません(図2)。植物はこれらの2つの機構により、季節変化などに起こる緩やかな温度低下と異常気象時や夜間などに起こる急激な温度低下を適切に感知して、効率的に耐性機構を働かせていると考えられます。
発表雑誌
- 雑誌名
- :「The Plant Cell」(3月28日オンライン版)
- 論文タイトル
- :Differential signaling in cold responses to rapid and gradual temperature decreases in Arabidopsis
- 著者
- :
Satoshi Kidokoro, Koshi Yoneda, Hironori Takasaki, Fuminori Takahashi, Kazuo Shinozaki, Kazuko Yamaguchi-Shinozaki*
- DOI番号
- :10.1038/nplants.2017.39
問い合わせ先
- 東京大学大学院農学生命科学研究科応用生命化学専攻 植物分子生理学研究室
- 教授 篠崎 和子(しのざき かずこ)
- TEL:03-5841-8137
- Fax:03-5841-8009
- E-mail:akys<アット>mail.ecc.u-tokyo.ac.jp <アット>を@に変えてください。
- 東京大学大学院農学生命科学研究科応用生命化学専攻 植物分子生理学研究室
- 助教 城所 聡(きどころ さとし)
- TEL:03-5841-8111
- Fax:03-5841-8009
- E-mail:akido<アット>mail.ecc.u-tokyo.ac.jp <アット>を@に変えてください。
用語解説
- 注1.転写因子
-
標的とする遺伝子のプロモーター配列(DNA)に結合し、その遺伝子の発現を活性化したり抑制したりして調節するDNA結合性タンパク質。DREB1は低温誘導性遺伝子発現の活性化因子として、CAMTAはDREB1BとDREB1C遺伝子発現の活性化因子として働いている。
- 注2.概日時計
-
1日を周期とした概日リズムを生み出す体内時計の分子機構で、LHYやCCA1、TOC1などと名付けられたタンパク質が中心振動子を構成している。
明暗サイクル(昼夜)などに同調してほぼ1日(24時間)周期で変動する応答パターン。