イネの防御物質生産における鍵遺伝子の発現を強化する因子の発見
〜有用フラボノイド生産に向けた応用にも期待〜
- 発表者
-
小川 哲史 (東京大学生物生産工学研究センター 大学院生)
宮本 皓司 (帝京大学理工学部 研究員)
根本 圭一郎 (愛媛大学プロテオサイエンスセンター 研究員)
澤崎 達也 (愛媛大学プロテオサイエンスセンター 教授)
山根 久和 (帝京大学理工学部 教授)
野尻 秀昭 (東京大学生物生産工学研究センター 教授)
岡田 憲典 (東京大学生物生産工学研究センター 准教授)
発表のポイント
◆イネの抗菌性フラボノイドであるサクラネチン(注1)の生合成では、ジャスモン酸(注2)シグナルのマスターレギュレーターである転写因子(注3)OsMYC2による制御が必須であることを明らかにしました。
◆OsMYC2が新規転写因子OsMYL1またはOsMYL2と相互作用することで活性化し、サクラネチン生合成遺伝子の発現を上昇させることを解明しました。
◆サクラネチンを高生産する病害抵抗性イネの作出、有用フラボノイドを生産する新たな作物の作出などへの応用が期待されます。
発表概要
植物は生存戦略の一環として様々な防御物質を生産しますが、その代表例がファイトアレキシンと呼ばれる抗菌性化合物です。イネにおいては17種類のファイトアレキシンが知られており、このうち唯一のフラボノイド型化合物であるサクラネチンは、抗菌活性の強さなどから主要なファイトアレキシンの一つとされています。しかし、その生産を調節する分子メカニズムは未解明のままでした。
東京大学生物生産工学研究センター岡田憲典准教授らの研究グループは、帝京大学バイオサイエンス学科、愛媛大学プロテオサイエンスセンターと共同で、サクラネチン生産の調節機構の解明を目的として研究を行い、特にサクラネチンの生産に必須である遺伝子OsNOMTの調節機構を詳細に解析しました。その結果、イネのジャスモン酸シグナルを広く調節する転写因子OsMYC2がOsNOMTの発現、ひいてはサクラネチンの生産において中心的な役割を担っていることを明らかにしました。さらに、OsMYC2が2つの新規転写因子OsMYC2-like 1, 2 (OsMYL1、OsMYL2) とタンパク質間相互作用を形成すること、その相互作用によりOsMYC2の転写活性化能が劇的に上昇することを示しました。
今後、OsMYC2、OsMYL1、OsMYL2による転写制御機構を活用することで、サクラネチンを高生産する病気に強いイネの育成や、ポリフェノールとして知られる有用フラボノイドの高生産作物の創出にもつながることが期待されます。
発表内容
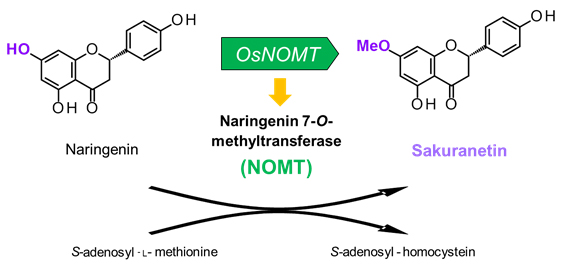
図1 サクラネチン生合成の最終ステップ
(拡大画像↗)
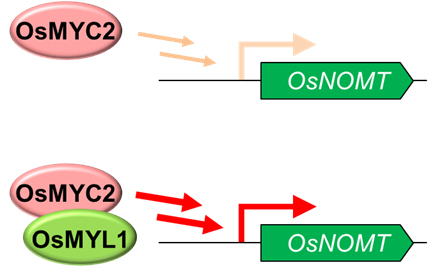
図2 OsMYL1あるいはOsMYL2とのタンパク質間相互作用によりOsMYC2の転写活性化能が増強される様子を示す模式図(図ではOsMYL1の場合を示す)。 (拡大画像↗)
〈研究の背景と目的〉
植物は病原菌から身を守るため、ジャスモン酸(JA)などを介したシグナル伝達によりファイトアレキシンと呼ばれる防御物質を生産します。イネにおいては、17種類のファイトアレキシンが知られており、このうち唯一のフラボノイド型化合物であるサクラネチンはいもち病菌感染時に多く蓄積し、強い抗菌活性を示すことから、主要なファイトアレキシンの一つとされています。サクラネチン生産はその誘導にJAを要求する防御応答であり、他の主要なファイトアレキシンの生産がJA要求性、非要求性いずれのシグナルでも誘導されることと対照的です。本研究では、JA要求性防御応答のモデルケースと考えられるサクラネチンの生産誘導に着目しました。
サクラネチン生合成の最終ステップでは、前駆体であるナリンゲニンが生合成酵素naringenin 7-O-methyltransferase (NOMT) によってサクラネチンに変換されます(図1)。先行研究において、このNOMTをコードする遺伝子OsNOMTが見出され (J. Biol. Chem. (2012): 2012/06/04プレスリリース参照) 、さらにOsNOMTの発現誘導はサクラネチンの生産誘導と同様にJAを要求する反応であることを明らかにしておりました。しかしながら、OsNOMTの発現機構の詳細については未解明のままでした。そこで、岡田准教授らは、JA要求性を示すサクラネチン生産の制御機構の解明を目的とし、特にOsNOMTの発現を制御するメカニズムの詳細な解析に取り組んできました。
〈研究の内容・意義〉
まず、OsNOMTの発現制御に関わる転写因子OsMYC2、およびOsMYC2と相同性のあるOsMYC2-like protein (OsMYL1) をトランスクリプトーム解析(注4)データから選抜した後、これらの機能解析によって、OsMYC2がOsNOMTのプロモーター活性を顕著に上昇させることを示しました。また、OsMYC2によるOsNOMTのプロモーター活性の上昇が、OsMYL1との共存により相乗的に亢進することを示しました。次に、OsMYC2の発現が抑制されたイネを作出しました。イネにおいてはOsMYC2が単一の遺伝子のためその欠損は致死となり、変異体の取得ができないと考えられました。そこで、OsMYC2の発現を様々なレベルで低下させられるRNAi法(注5)を用いてOsMYC2抑制株の作出を行いました。得られた抑制株においては、JA誘導的なOsNOMT発現およびサクラネチン蓄積と前駆体であるナリンゲニンの蓄積が抑制されることを明らかにし、OsNOMTの発現およびサクラネチンの生産がOsMYC2の制御下で起こっていることを証明しました。また、OsMYL1と相同性の高いOsMYL2についても、その転写活性化能をレポータージーンアッセイ(注6)により解析したところ、OsMYC2との共存下において相乗的なOsNOMTのプロモーター活性の上昇が認められました。これらの転写因子については、物理的なタンパク質間相互作用が生体内と試験管内の両方で起こることを実証し、さらにこの相互作用によってOsMYC2のもつ転写活性化能が大きく増強されることを明らかにしました。この一連の結果から、OsMYC2がOsMYL1およびOsMYL2と相互作用することで活性化し、OsNOMTの発現をJA誘導的に上昇させることで、サクラネチンの生産を促進する制御モデルを提唱しました。
〈今後の予定・期待〉
OsMYC2、OsMYL1、OsMYL2がどのような形で相互作用し、どのような機構でターゲット遺伝子の転写制御を可能にしているのかのメカニズムを解き明かす研究を進める予定です。今回、OsMYL1とOsMYL2は同様の機能を示しましたが、OsMYL1の働きがJAにより誘導されるのに対しOsMYL2は誘導されないという明確な差があります。したがって、植物体内においてOsMYC2-OsMYL1複合体とOsMYC2-OsMYL2複合体を状況により使い分けていることが考えられるため、OsNOMTの発現制御機構の解明においてこの使い分けを理解することも重要と考えられます。今後の進展により、将来的にはサクラネチンを高生産し強い病害抵抗性を示すイネの作出や有用フラボノイドを高生産する新たな作物の創出が期待されます。
発表雑誌
- 雑誌名
- :Scientific Reports
- 論文タイトル
- :OsMYC2, an essential factor for JA-inductive sakuranetin production in rice, interacts with MYC2-like proteins that enhance its transactivation ability
- 著者
- :Satoshi Ogawa, Koji Miyamoto, Keiichirou Nemoto, Tatsuya Sawasaki, Hisakazu Yamane, Hideaki Nojiri, and Kazunori Okada
- DOI番号
- :10.1038/srep40175
- 論文URL
- :http://www.nature.com/articles/srep40175
問い合わせ先
- 東京大学生物生産工学研究センター 環境保全工学研究室
准教授 岡田 憲典 (おかだ かずのり) - Tel:03-5841-3070
Fax:03-5841-8030 - Email: ukazokad@mail.ecc.u-tokyo.ac.jp
研究室URL:http://park.itc.u-tokyo.ac.jp/biotec-res-ctr/kampo/index.html
用語解説
- (注1)サクラネチン
- イネが生産するフラボノイド型化合物であり、ファイトアレキシンと呼ばれる抗菌性化合物の一種。いもち病菌に対し、胞子の発芽阻害活性や菌糸の伸長阻害活性を強く示す。サクラネチンはイネのファイトアレキシンとしてのみならず、抗炎症効果や抗病原活性を有するなど医薬品としても有用な化合物であると期待されている。
- (注2)ジャスモン酸(JA)
- 植物ホルモンの一種。メチルエステルであるジャスモン酸メチルはジャスミンの主要な香気成分である。ジャスモン酸が植物にもたらす生理作用は、防御応答の誘導や形態形成の調節など多岐に及ぶ。
- (注3)転写因子
- その名の通り、遺伝子の転写を調節するタンパク質。標的とする遺伝子の周辺に存在する特定のDNA配列に作用することで遺伝子の働きを強化あるいは抑制し(その強さを転写活性化能および転写抑制能と呼ぶ)、最終的に防御応答や成長に影響を与える。転写因子の調節機能は、他のタンパク質との相互作用やリン酸化などの修飾によって変化することがある。
- (注4)トランスクリプトーム解析
- ある条件下における、細胞内すべての遺伝子の発現量を調べる手法(transcript; 転写 + -ome; 全)。これにより、例えばストレスを与えた際に働く遺伝子群を決定することができる。
- (注5)RNAi法
- RNA interferenceの略、日本語でRNA干渉とも呼ばれる遺伝子の発現抑制の方法。二本鎖RNAと相補的な塩基配列を持つmRNAが分解される現象を利用して、人工的に二本鎖RNAを植物体内で合成させることにより、任意の遺伝子の発現を抑制できる。ゲノム編集によるノックアウトと異なり、様々な発現抑制レベルの個体を得ることができるため、今回の様に致死遺伝子の抑制の場合にも有効な場合がある。
- (注6)レポータージーンアッセイ
- あるDNA配列が転写に与える影響を調べる方法。蛍光タンパク質などをコードするレポーター遺伝子に標的のDNA配列を連結し、細胞内におけるレポーターの活性を調べることで影響を評価する。この際、エフェクターと呼ばれる因子を同時に細胞内に導入することで、エフェクターが転写に与える影響を評価することもできる。