小胞体からミトコンドリアへステロールが輸送される機構を解明
- 発表者
-
田 スチ(東京大学大学院農学生命科学研究科応用生命工学専攻 博士課程(当時)、
福田 良一(東京大学大学院農学生命科学研究科 応用生命工学専攻 助教)
日本学術振興会特別研究員 DC-2(当時))
太田 明徳(中部大学応用生物学部応用生物化学科 教授)
堀内 裕之(東京大学大学院農学生命科学研究科 応用生命工学専攻 教授)
発表のポイント
◆ORPは酵母から動物、植物まで真核生物に広く保存されていますが、今回の発見によりORPの新たな生理機能が明らかになりました。
◆細胞内のステロール輸送機構の解明は、真核細胞の細胞小器官の形成機構の理解に貢献するとともに、ORP遺伝子の変異に起因する代謝関連疾患などの発症機構の解明につながることが期待されます。
発表概要
東京大学大学院農学生命科学研究科の福田良一助教、堀内裕之教授らは、酵母においてオキシステロール結合タンパク質(Oxysterol-binding protein, OSBP)関連タンパク質(OSBP Related Protein, ORP)が小胞体からミトコンドリアへのステロールの輸送を担うことを明らかにしました。小胞体で新規合成されたステロールの各オルガネラへの輸送機構はオルガネラの形成に関わる細胞生物学の基本的かつ重要な課題であり、本研究の成果は真核細胞の細胞構造形成の基礎理解に貢献するものです。また、ヒトでは一部のORP遺伝子の変異と脂質代謝関連疾患などと関わりが報告されていることから、本研究の成果はそれらの疾患の発症機構の理解にも寄与すると期待されます。
発表内容
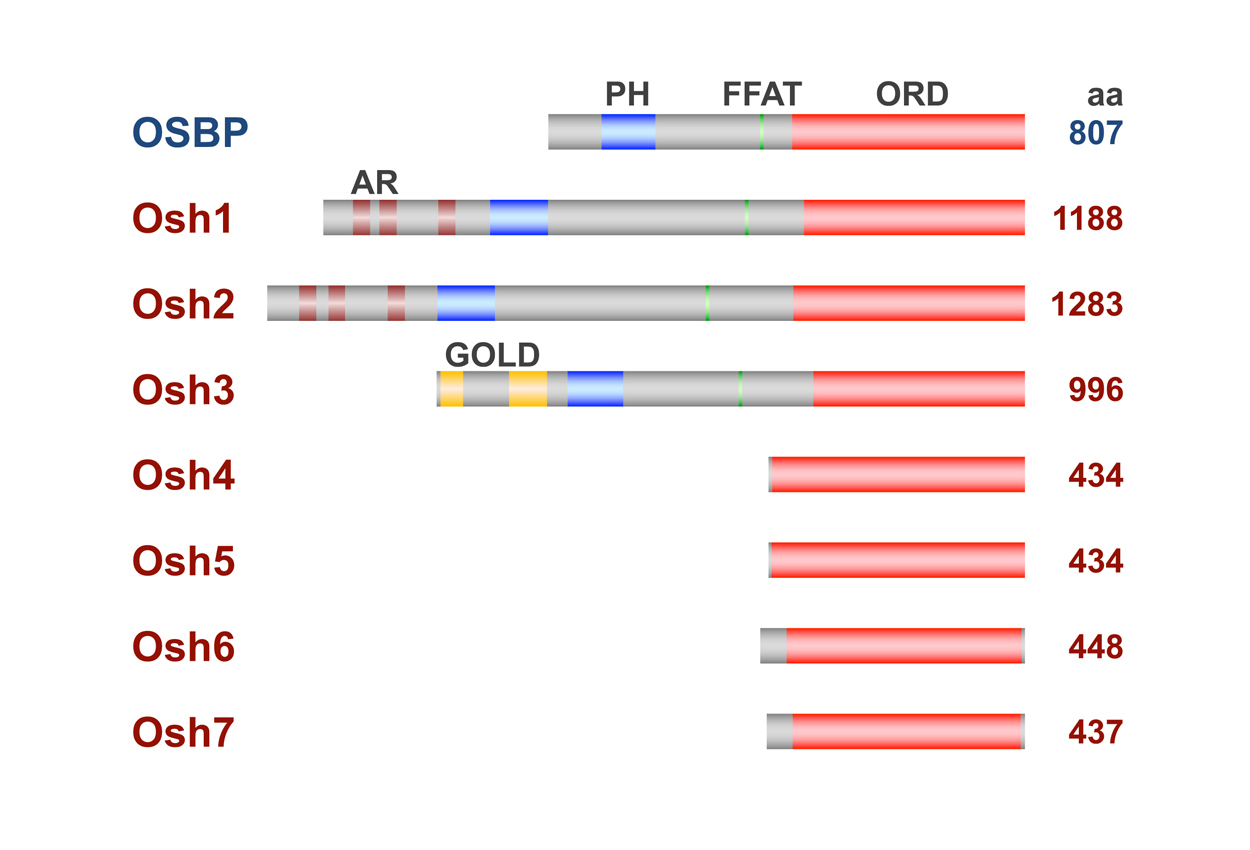
図1 ヒトのOSBPとS. cerevisiaeのORPの構造
S. cerevisiaeは7種のORP、Osh1〜Osh7を有する。ORPはリガンドとの結合に関わるOSBP-related domain(ORD)を共通して持つ。また、一部のORPにはホスホイノシチドとの結合に関わるPH domainや小胞体膜タンパク質であるVAPとの結合に関わるFFATモチーフ、タンパク質との結合に関わるアンキリンリピート(AR)、ゴルジ体タンパク質に見られるGoldドメインなどが存在する。 (拡大画像↗)
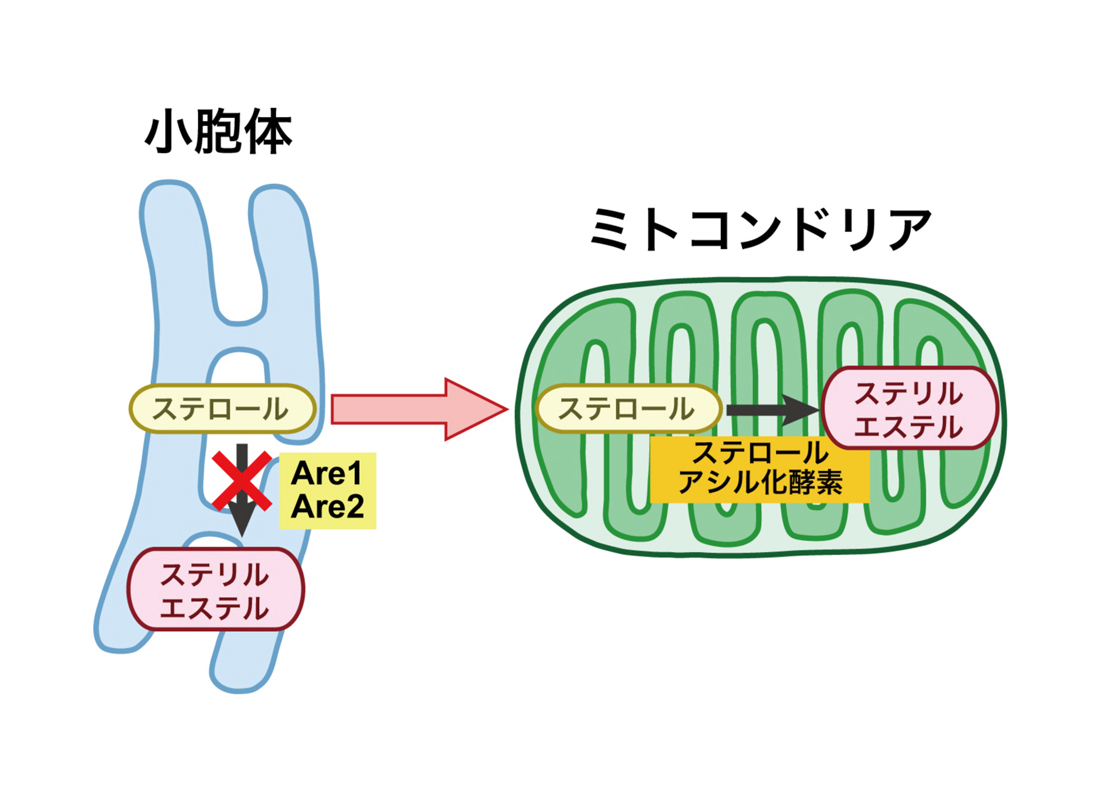
図2 小胞体からミトコンドリアへのステロール輸送の解析
酵母のアシルCoA:ステロールアシル化酵素遺伝子ARE1およびARE2を破壊し、ミトコンドリアに異種のステロールアシル化酵素を局在させた株において、ミトコンドリアでステロールがエステル化されることを指標にステロール輸送を解析した。 (拡大画像↗)
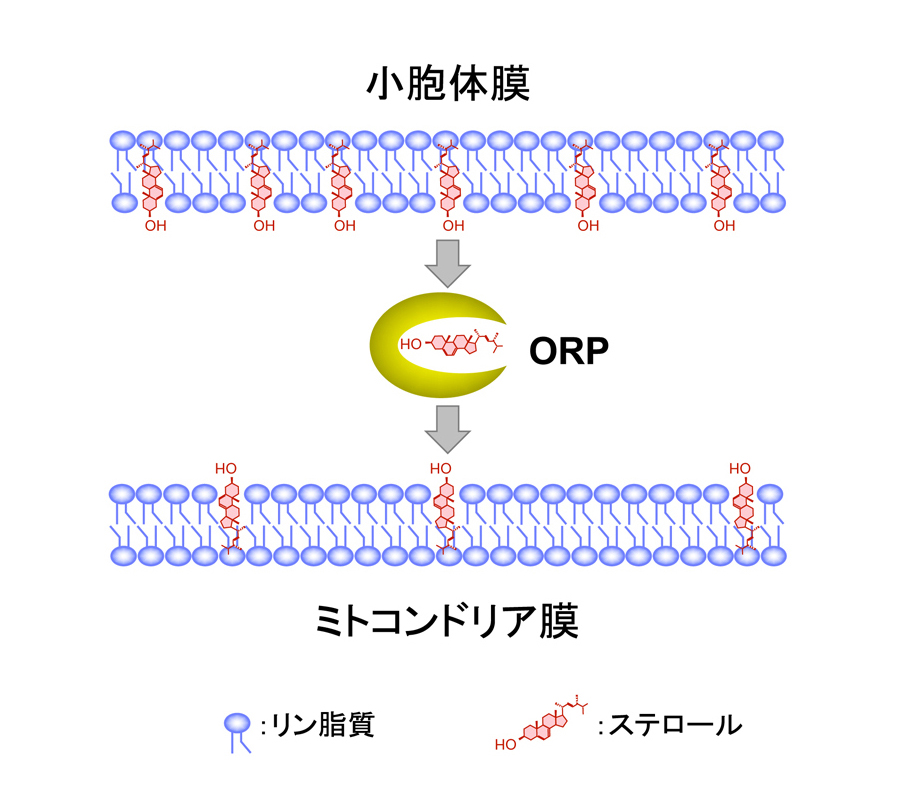
図3 ORPによる小胞体からミトコンドリアへのステロール輸送
ORPは小胞体膜上で合成されたステロールを引き抜き、ミトコンドリア膜に挿入することによりステロールを輸送する。 (拡大画像↗)
ステロールは真核細胞の生体膜の構成成分の1つであり、生存に必須の脂質です。ステロールは膜の流動性や透過性に関与するとともに、膜上にマイクロドメインを形成して情報伝達などに関わると考えられています。ステロールは細胞膜やほとんどの細胞小器官(オルガネラ)の膜に様々な割合で含まれていますが、一方で細胞はステロールを小胞体で新規合成するか細胞外から取り込むことによって獲得し、それが必要に応じてそれらの膜へ分配されています。ステロールはミトコンドリアの膜にも含まれています。酵母Saccharomyces cerevisiaeではミトコンドリアはチューブ状の構造を取りますが、主要ステロールであるエルゴステロールが欠乏するとミトコンドリアは凝集してしまうことから、ステロールはミトコンドリアの構造の維持に重要な役割を果たしていると考えられています。また、動物では、主要ステロールであるコレステロールはミトコンドリアでステロイドホルモンなどに変換されます。従って、生物にとって小胞体で合成したステロールをミトコンドリアへ適切に輸送することは極めて重要であると考えられます。しかしながら、小胞体からミトコンドリアへどの様にステロールが輸送されるかは酵母から動物、植物に至るまで未解明でした。
細胞内の脂質輸送機構の一つとして、脂質分子を一方の膜から引き抜き、もう一方の膜へ輸送し挿入する脂質輸送タンパク質(Lipid Transfer Protein, LTP)による脂質輸送が想定されています。オキシステロール結合タンパク質(Oxysterol-binding protein, OSBP)はもともと動物において酸化コレステロールと結合するタンパク質として見出されましたが、OSBPと相同性を持つOSBP関連タンパク質(OSBP related protein, ORP)は真核生物に広く保存されており、ヒトは12種のORP遺伝子を、S. cerevisiaeはOSH1〜OSH7という7種のORP遺伝子を持っています(図1)。ORPの機能としては、細胞内のステロール輸送を担うLTPとして機能する可能性が考えられてきましたが、一方でORPが脂質センサーあるいは制御因子として小胞輸送やリン脂質代謝、シグナル伝達など様々な細胞内プロセスに関与するということが報告されており、ORPの機能については明確な結論が得られていない状況でした。S. cerevisiaeでは、OSH1〜OSH7を単独で破壊した株は生育はできますが、全て破壊すると致死となることから、Osh1〜Osh7は生育に必須の機能を共有していると考えられていますが、それが何かは不明です。
東京大学大学院農学生命科学研究科の福田良一助教、堀内裕之教授らの研究グループは、S. cerevisiaeから精製した小胞体およびミトコンドリアを用いてステロール輸送をin vitroで解析する系を新たに構築し、その機構を解析しました。通常細胞内で合成されたステロールは細胞膜やオルガネラに輸送される以外に、一部は小胞体膜上のアシルCoA:ステロールアシル化酵素によってステリルエステルに変換され、脂肪滴に貯蔵されます。酵母においてアシルCoA:ステロールアシル化酵素をコードするARE1とARE2を破壊した株では、ステリルエステルが検出されなくなりますが、生育には大きな影響は見られません。そこで、この株において細菌Aeromonas salmonicida由来のステロールアシル化酵素を人為的にミトコンドリアに局在させ、ステロールがミトコンドリアに輸送されてエステル化されることを指標に、ステロールの輸送を解析しました(図2)。その結果、酵母から単離した小胞体、ミトコンドリア、細胞質を用いてin vitroでのステロール輸送反応を行った場合には小胞体からミトコンドリアへのステロール輸送が観察されましたが、Oshタンパク質を全て欠損させた場合にはステロール輸送が低下しました。さらに細胞質画分の代わりに、精製したOsh4〜Osh7タンパク質を添加した場合や、Osh1〜Osh3を単独で発現する株から調製した細胞質画分を添加した場合にはステロール輸送が回復しました。また、OSH遺伝子を全て欠損させた株ではミトコンドリアのステロール含量が低下しました。これらの結果から、小胞体からミトコンドリアへORPによってステロールが輸送されることが明らかになり(図3)、酵母のORPは全てステロールを輸送する機能を持つことが示唆されました。
本研究により、これまで謎であった小胞体からミトコンドリアへのステロールの輸送機構の一端が初めて明らかにされました。小胞体で合成されたステロールの各オルガネラへの輸送機構は、真核細胞の基本構造であるオルガネラ膜の形成に関わる細胞生物学の重要な課題であり、本研究の成果は真核細胞の細胞構造形成の基礎理解に大きく貢献するものであると考えられます。また、本研究によりORPの新たな機能が明らかになっただけでなく、酵母のORPが全てステロールを輸送する機能を備えていたことから、ORPの必須の機能はステロールを輸送することである可能性が考えられました。ヒトでは、一部のORP遺伝子の変異と脂質代謝関連疾患などとの関わりが報告されており、本研究で得られたORPの基本機能に関する知見はそれらの疾患の発症機構の理解にも貢献するものであると期待されます。
この研究は、日本学術振興会科学研究費補助金(17688005、20580072、16J00457)、公益財団法人アサヒグループ学術振興財団研究助成、公益財団法人エリザベス・アーノルド富士財団研究助成の支援を受けて行われました。発表雑誌
- 雑誌名
- :The Journal of Biological Chemistry(293巻、(2018年)、5636〜5648ページ)
- 論文タイトル
- :Oxysterol-binding protein homologs mediate sterol transport from the endoplasmic reticulum to mitochondria in yeast
- 著者
- :Siqi Tian1, Akinori Ohta2, Hiroyuki Horiuchi1, and Ryouichi Fukuda1*
1 東京大学大学院農学生命科学研究科 応用生命工学専攻
2 中部大学応用生物学部応用生物化学科 - DOI番号
- :10.1074/jbc.RA117.000596
- 論文URL
- :http://www.jbc.org/content/293/15/5636.short
問い合わせ先
助教 福田 良一(ふくだ りょういち)
Tel:03-5841-5178
E-mail:afukuda<アット>mail.ecc.u-tokyo.ac.jp <アット>を@に変えてください。
教授 堀内 裕之(ほりうち ひろゆき)
Tel:03-5841-5169
E-mail:ahhoriu<アット>mail.ecc.u-tokyo.ac.jp <アット>を@に変えてください。
研究室URL:http://park.itc.u-tokyo.ac.jp/cell-gene/index.html
用語解説
- 注1 ステロール
- シクロペンタノヒドロフェナントレン環を持ち、3位に水酸基を有する脂質。広く真核細胞に存在し、生体膜を構成する。ステロールはステロイドホルモンや胆汁酸、ビタミンの合成中間体でもある。ヒトを含む動物ではコレステロールが主要ステロールであるのに対して、酵母などの菌類ではエルゴステロールが主要ステロールである。
- 注2 Saccharomyces cerevisiae
- 古くからアルコール醸造やパン作りに用いられてきた酵母。真核細胞のモデル生物として生命科学研究に利用されている。