「魔女の雑草」の発芽誘導物質に対する選択性を原子レベルで解明
~根寄生雑草防除剤の合理的デザインに貢献~
- 発表者
-
徐 玉群(東京大学大学院農学生命科学研究科 応用生命化学専攻 特任助教)
宮川 拓也(東京大学大学院農学生命科学研究科 応用生命化学専攻 特任准教授)
野﨑 翔平(東京大学大学院農学生命科学研究科 応用生命化学専攻 博士課程3年生)
中村 顕(東京大学大学院農学生命科学研究科 応用生命化学専攻 特任助教:研究当時)
呂 瑩(東京大学大学院農学生命科学研究科 応用生命化学専攻 修士課程学生:研究当時)
中村 英光(東京大学大学院農学生命科学研究科 応用生命化学専攻 助教)
大戸 梅治(東京大学大学院薬学系研究科 准教授)
石田 英子(東京大学大学院薬学系研究科 特任研究員)
清水 敏之(東京大学大学院薬学系研究科 教授)
浅見 忠男(東京大学大学院農学生命科学研究科 応用生命化学専攻 教授)
田之倉 優(東京大学大学院農学生命科学研究科 応用生命化学専攻 特任教授)
発表のポイント
◆ストライガの3つのクレード(注3)に属するHTLタンパク質とD14タンパク質が、高いアミノ酸配列の相同性にも関わらず発芽誘導物質に対する選択性が異なる理由を、構造の比較解析により説明しました。
◆本研究で見出された構造基盤は、ストライガのHTL/D14を標的としたストライガ防除剤を合理的にデザインする上でも有用な知見であると考えられます。
発表概要
東京大学大学院農学生命科学研究科の田之倉優特任教授と浅見忠男教授を中心とした研究グループは、SLに高感受性の受容体ShHTL4及びShHTL7(多様型クレード)、低感受性のShHTL1(保存型クレード)及びShD14のリガンド(注5)選択性を解析し、それらの立体構造をX線結晶構造解析法(注6)により決定しました。当グループが以前に報告したShHTL3(中間型クレード)の立体構造を併せて比較解析することにより、ストライガ特異的に3つのクレードに属するShHTLとShD14が異なるリガンド選択性をもつ理由を詳細に説明することができました。さらに、下流のシグナル伝達因子との相互作用を解析することで、ShHTL及びShD14を介するシグナル伝達経路がリガンド選択性により異なる分子モデルを提案しました。本研究で明らかにした構造基盤は、ストライガが寄生シグナル物質のSLを受容できるように至るまでのHTL/D14ファミリーの進化的な考察を提供するだけにとどまらず、ShHTLを標的としたストライガの自殺発芽誘導剤をデザインする上で役立つ知見になると考えられます。
発表内容

図1 ShHTL及びShD14の構造比較
3つのクレードに属するShHTLとShD14の全体構造を比較するすると、リガンド結合ポケットの入り口を構成しているαD1ヘリックスに構造の違いが見られました。特に多様型クレードのShHTL7(紫色)と中間型クレードのShHTL3(青色)のαD1ヘリックスに、最大5 Åの大きなずれが見られました。 (拡大画像↗)
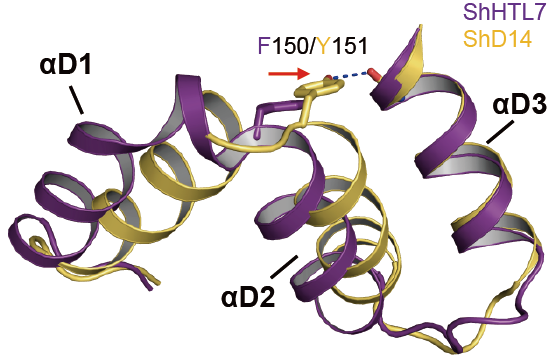
図2 ShHTL7とShD14の構造の重ね合わせ(一部)
保存型及び中間型クレードのShHTLとShD14の150/151番目のチロシン残基(Y150/Y151)によりαD1ヘリックスとαD3ヘリックスとの間に水素結合が形成されていましたが、多様型クレードのShHTLにおいてはチロシン残基がフェニルアラニン残基(F150)に置き換わっており水素結合がなくなっていました。 (拡大画像↗)
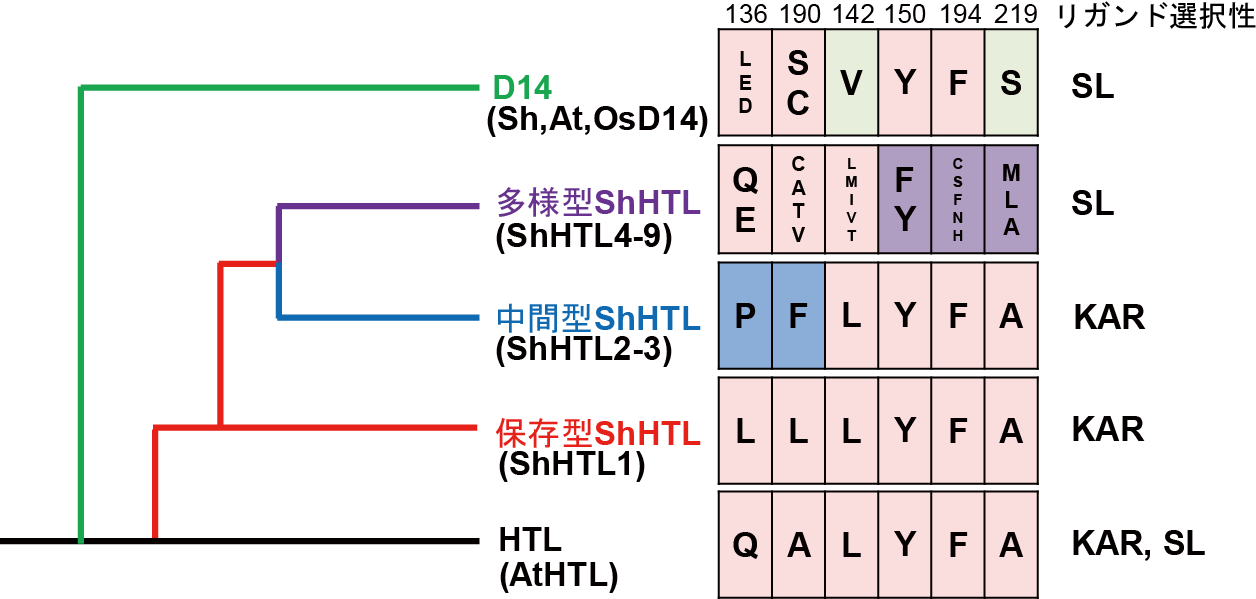
図3 ShHTLとShD14の異なるリガンド選択性の進化モデル
HTL/D14ファミリーの分子系統樹(左図)と進化上重要なアミノ酸残基及びそれぞれの認識するリガンド(右図)を示しています。多様型クレードに属するShHTLは、リガンド結合ポケットに側鎖が比較的小さな残基(C, S, Tなど)をもつ一方、保存型及び中間型ShHTLのポケットは嵩高い残基(Y, Fなど)で構成されています。また、142番目の残基とその配置を決める150番目の残基はSLとの結合に、219番目のアミノ酸残基はKARとの結合に重要です。保存型と中間型ShHTL及びシロイヌナズナのHTLタンパク質(AtHTL)の142番目の残基はLとなり、SLと立体障害が生じるため、SLとは結合できません。D14において対応する残基は側鎖が小さなVであり、多様型ShHTLはhelix αD1の配向がずれている影響で142番目の残基による立体障害が起こりません。同様に、保存型と中間型ShHTLの219番目のアミノ酸残基はAであり、KARと立体障害なく結合できます。このようなアミノ酸残基の違いにより、ShHTL及びShD14のリガンド特異性が生み出されると考えられます。 (拡大画像↗)
これまでイネやシロイヌナズナ由来のHTL及びD14やストライガ由来の中間型及び多様型クレードShHTLを含むHTL/D14ファミリーの立体構造はいくつか報告されていましたが、ストライガに存在するShHTL及びShD14のアミノ酸配列とリガンド選択性を明確に結びつけることはできていませんでした。そこで本研究では、SL高感受性のShHTL7を含む2種の多様型クレードShHTL(ShHTL7及びShHTL4)、保存型クレードShHTL1及びShD14の立体構造をX線結晶構造解析法により決定し、これまで報告されたHTL/D14ファミリーの立体構造と併せることで網羅的な比較解析を試みました。
まず、KAR結合能とSL加水分解活性を解析し、3つのクレードに属するShHTLとShD14のリガンド選択性をタンパク質レベルで初めて明らかにしました。等温滴定型熱量測定(注8)により保存型及び中間型クレードのShHTLがシロイヌナズナのHTLタンパク質と同程度のKAR結合能をもつことが示され、一方で多様型クレードのShHTLとShD14はSLの加水分解活性をもつことが示されました。これらの結果から、保存型及び中間型クレードのShHTLはKAR受容体として、多様型クレードのShHTLとShD14はSL受容体として機能しうると考えられます。
次に、ShHTLとShD14の立体構造をX線結晶構造解析により決定し、全体構造が類似していることが示されました。しかしながら、リガンド結合ポケットの入り口を構成するαD1へリックス(注9)とαD2ヘリックスの配向に明確な構造の違いがあることがわかりました(図1)。モデル植物のHTL/D14ファミリーとストライガの保存型及び中間型クレードShHTLに対し、多様型クレードShHTLにおいては、αD1ヘリックスがαD3へリックスから離れるように位置していました。前者においては150番目のチロシン残基によりαD1ヘリックスとαD3ヘリックスとの間の水素結合が形成されていましたが、後者においてはチロシン残基がフェニルアラニン残基に置き換わっており水素結合が消失していました(図2)。この結果、SLに高感受性である多様型クレードShHTLは、自身のαD1ヘリックスをαD3へリックスから遠ざけることにより、基質のポケットの入り口を大きくしていることがわかりました。さらにリガンド結合ポケットの内側を構成する残基を比較すると、モデル植物のHTLタンパク質、保存型及び中間型クレードShHTLでは嵩高い残基をもつ一方、多様型クレードShHTLではスレオニン残基やシステイン残基のような側鎖が比較的小さな残基をもつことがわかりました(図3)。以上のような各クレード特異的なアミノ酸残基の違いにより、KARのような小さな分子を認識するShHTLはリガンド結合ポケットが小さく、多様なサイズのSLを認識できるShHTLほどポケットが大きくなっていました。本研究において見出された構造基盤は、αD1ヘリックスの配向とポケット内部の残基の嵩高さにより規定されるポケットサイズがリガンド特異性と密接な関係にあることを強く示唆しています。
さらに本研究グループは、イネやシロイヌナズナにおいてF-boxタンパク質(注10)MAX2がSL及びKAR応答の両方に不可欠であることが知られていることから、ストライガにおけるMAX2オルソログのShMAX2に注目し、ShHTL及びShD14のリガンド特異性とShMAX2との相互作用の関連を解析しました。その結果、多様型クレードのShHTLとShD14はSL依存的にShMAX2と相互作用することが示されました。両者はそれぞれストライガにおける外来性SLと内在性SLの受容体として機能する可能性が考えられます。また、ShHTLとシロイヌナズナのHTLタンパク質はKAR依存的にMAX2タンパク質と直接相互作用しないことが示され、KARはSLとは異なるメカニズムにより発芽誘導を活性化することが示唆されました。
本研究は、ストライガの発芽誘導に関わるShHTLのリガンド選択性とそれを規定する構造基盤を解明し、SLによるストライガ発芽誘導の分子メカニズムの一端を明らかにしました。また、解明したリガンド選択性の構造基盤は根寄生植物の発芽を制御する化合物の設計等に役立つと期待されます。
発表雑誌
- 雑誌名
- : 「Nature Communications」(平成30年9月26日)
- 論文タイトル
- :Structural analysis of HTL and D14 proteins reveals the basis for ligand selectively in Striga
(ストライガにおけるHTL及びD14タンパク質の構造解析によって明らかになったリガンド選択性の構造基盤) - 著者
- :Yuqun Xu, Takuya Miyakawa, Shohei Nosaki, Akira Nakamura, Ying Lyu, Hidemitsu Nakamura, Umeharu Ohto, Hanako Ishida, Toshiyuki Shimizu, Tadao Asami, and Masaru Tanokura*
- DOI番号
- :10.1038/s41467-018-06452-2
- 論文URL
- :https://www.nature.com/articles/s41467-018-06452-2
問い合わせ先
特任准教授 宮川 拓也(みやかわ たくや)
Tel:03-5841-2283
Faxl:03-5841-2283
E-mail:atmiya<アット>mail.ecc.u-tokyo.ac.jp <アット>を@に変えてください。
東京大学大学院農学生命科学研究科 応用生命化学専攻 養生訓を科学する医食農連携寄付講座
特任教授 田之倉 優(たのくら まさる)
Tel:03-5841-5165
Faxl:03-5841-8023
E-mail:amtanok<アット>mail.ecc.u-tokyo.ac.jp <アット>を@に変えてください。
用語解説
- 注1 根寄生植物ストライガ
- 根寄生植物は他の植物の根に寄生し栄養分を吸収して生育する植物です。特に、ストライガ属とオロバンキ属の根寄生植物は主要な作物に寄生し、サハラ以南のアフリカの農業生産に甚大な被害を与えています。
- 注2 HTL/D14ファミリー
- HTLタンパク質はカリキンの候補受容体、D14タンパク質はストリゴラクトンの受容体として知られています。HTLとD14の構造は非常に類似しており、HTL/D14ファミリーとして分類されます。近年、寄生植物ストライガのHTL遺伝子は独自に進化し、非寄生植物シロイヌナズナのD14遺伝子と同様にストリゴラクトンを受容する機能を獲得したと指摘されています。
- 注3 クレード
- 分子系統樹上で進化距離により分けられたグループ又は分岐群のことです。同じクレードの遺伝子は高いアミノ酸配列の相同性をもちます。
- 注4 ストリゴラクトン(SL)
- ストライガの発芽誘導活性をもつ物質として最初に発見された化合物です。ストリゴラクトンは、植物の枝分かれを抑制する植物ホルモンとして、また菌根菌を呼び寄せて共生するために植物の根より分泌される物質として機能することも知られています。
- 注5 リガンド
- 特定のタンパク質(受容体と呼ばれる)に特異的に結合する物質のことです。多くの場合、リガンドは受容体の特定の部位に高い親和性で結合することで、受容体の機能を調節します。
- 注6 X線結晶構造解析法
- 解析したいタンパク質を高純度に精製し、適した溶液組成・温度等の条件下で徐々に析出させていくと、分子が規則正しく並んだ結晶が形成します。この結晶にX線を照射して得られるX線回折像を解析することで、タンパク質の三次元構造を決定することができます。
- 注7 カリキン(KAR)
- 山火事などで植物が燃焼した際に発生する煙に含まれ、発芽誘導をもつブテノリド類の物質です。現在、6種のカリキン類が知られています。
- 注8 等温滴定型熱量測定
- 一方の分子の溶液を標的分子に滴下した際に起こる化学反応もしくは結合反応に伴う熱変化(吸熱もしくは発熱)を観測する熱力学的測定法です。この方法では、相互作用の結合定数、反応の結合比、エンタルピー変化及びエントロピー変化などの熱力学的パラメータが得られるため、分子間相互作用の解析によく利用されています。
- 注9 αヘリックス
- タンパク質に見られる特徴的な二次構造の一つであり、ペプチド鎖が右巻きのらせん構造をとっています。
- 注10 F-boxタンパク質
- F-boxモチーフを含むタンパク質です。F-boxタンパク質は、Skp1タンパク質、Cullinタンパク質及びRBX1タンパク質とE3ユビキチンリガーゼ複合体を形成します。E3ユビキチンリガーゼ複合体によりユビキチン標識されたタンパク質はプロテアソームによって分解されます。