ハワイのキハダは空気から
-亜熱帯外洋域魚類生産における窒素固定生物の寄与を解明-
- 発表者
- 高橋 一生(東京大学大学院農学生命科学研究科 准教授)
堀井 幸子(東京大学大学院農学生命科学研究科 博士課程3年・日本学術振興会特別研究員(DC1))
塩崎 拓平(海洋研究開発機構 特任研究員)
橋濱 史典(東京海洋大学学術研究院 助教)
古谷 研(研究当時:東京大学大学院農学生命科学研究科 教授、現:創価大学)
発表のポイント
◆窒素ガスから出発する食物連鎖がキハダマグロの生産に直接つながっていることを初めて明らかにした。
◆従来深層から栄養供給で魚類生産は支えられていると考えられてきたが「海の砂漠」とよばれる栄養の乏しい亜熱帯外洋域では、窒素ガス由来の栄養が重要であることを示し、海洋の新たな生物生産メカニズムを明らかにした。
発表概要
全海洋のおよそ6割を占める亜熱帯外洋域表層は栄養塩が枯渇しているため、植物プランクトンの現存量が低く、「海の砂漠」と呼ばれてきました。近年、この海域では、窒素ガスを栄養源として利用できる窒素固定者が基礎生産者として重要であると考えられるようになってきましたが、その生産物が魚類生産にまで転送されているかどうかについては明らかにされてきませんでした。東京大学大学院農学生命科学研究科、海洋研究開発機構、東京海洋大学らの研究グループは、中央太平洋において、現場窒素固定速度、硝酸塩濃度および、食物連鎖構成各要素の炭素窒素安定同位体比を測定し、基礎生産者が利用している栄養源(窒素)と、その生産物が食物連鎖で上位栄養段階に転送される過程について調査を行いました。その結果、太平洋外洋亜熱帯域には、窒素固定者の寄与が明瞭に異なる三つの食物連鎖が存在していることが明らかとなりました。このうちハワイ周辺を含む15〜25°Nおよび20〜30°Sにおける食物連鎖はほぼ窒素固定者により支えていると考えられ、例えばハワイ周辺で漁獲されたキハダマグロのタンパク質生産には空気由来の窒素が寄与していることが示されました。今回の研究結果は、一見生産性の低い亜熱帯外洋域が、なぜカツオ、マグロ類などを始めとする重要な水産資源の漁場となっているのか、そのメカニズムを考える上で重要な成果であると言えます。
発表内容
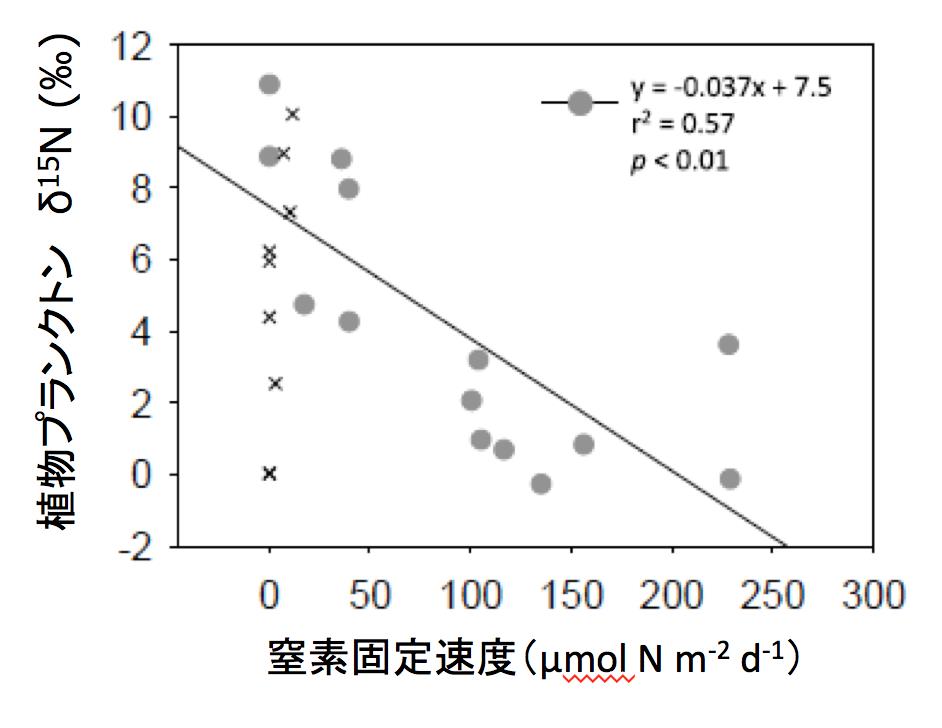
図1 中央太平洋南北縦断測線上測点表層における窒素固定速度と植物プランクトン(POM)窒素安定同位体比の関係. ●:亜熱帯海域、×:その他の海域(拡大画像↗)
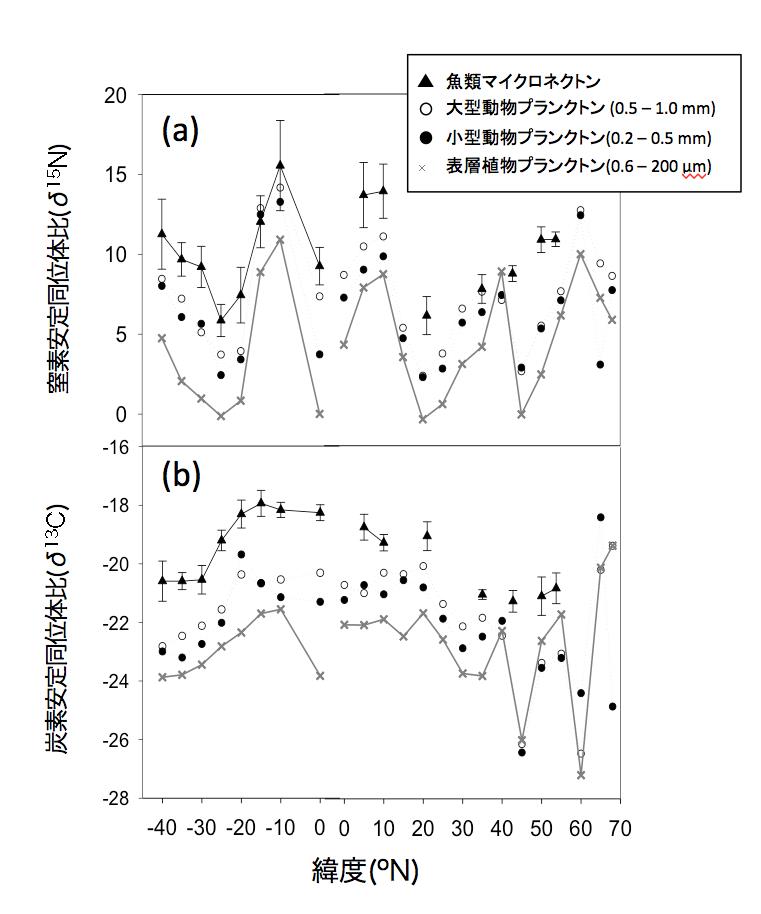
図2 中央太平洋南北縦断測線上測点における、植物プランクトン(POM)、動物プランクトン、および魚類マイクロネクトンの安定同位体比変動. (a) 窒素、(b)炭素(拡大画像↗)
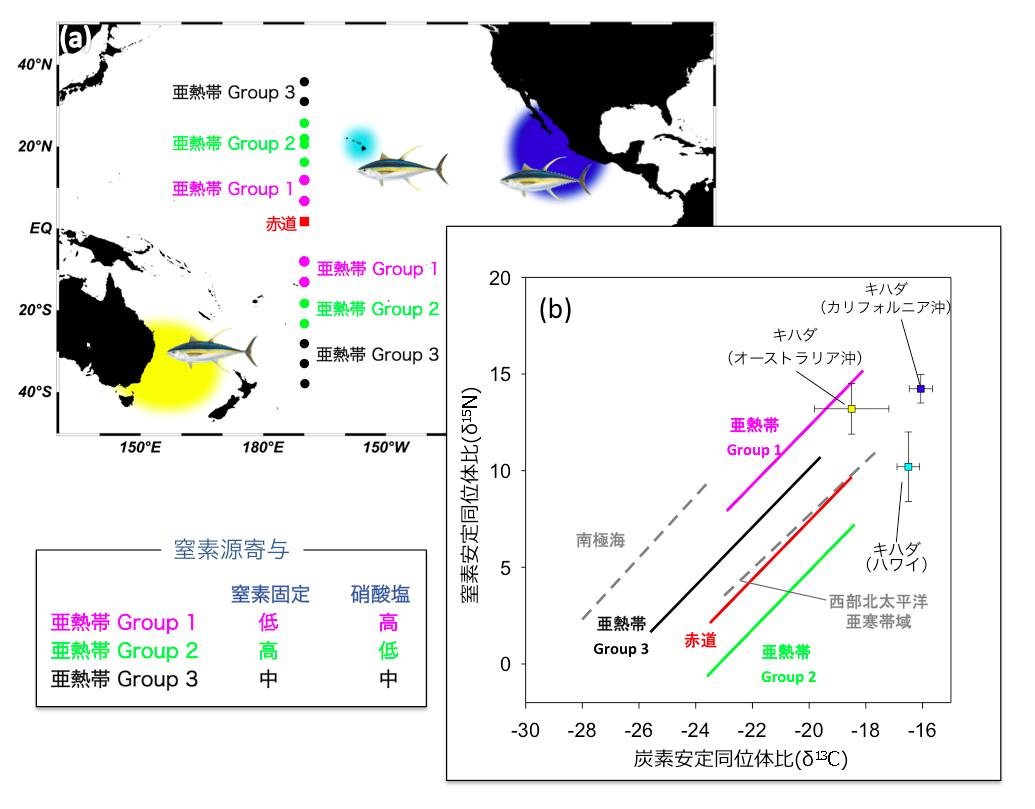
図3 中央太平洋観測線上測点表層より得られた食物連鎖構成要素(植物プランクトン、動物プランクトン、魚類マイクロネクトン)の炭素窒素安定同位体比の特徴に基づいて区分された(a)各測点の地理的配置および解析に用いた高次捕食者(キハダマグロ)の採集海域(既報研究)、と(b)食物連鎖構造の特徴と高次捕食者(キハダマグロ)の安定同位体比との関係.
太平洋他海域(南極海、北太平洋亜寒帯域)より得られた安定同位体比に基づく食物連鎖構造の関係(既往研究)も同時に示してある。窒素固定が安定的に優勢する海域(Group 2)の食物網は、窒素固定のほとんどない海域(Group 1)や変動の大きい海域(Group 3)と、地理的にも同位体比の特徴からも明確に区分されることがわかる。またハワイのキハダマグロの安定同位体比のプロットは、窒素固定優勢海域(Group 2)ラインの延長線上に位置していることから、その生産が窒素固定者に支えられていることが示唆される。(拡大画像↗)
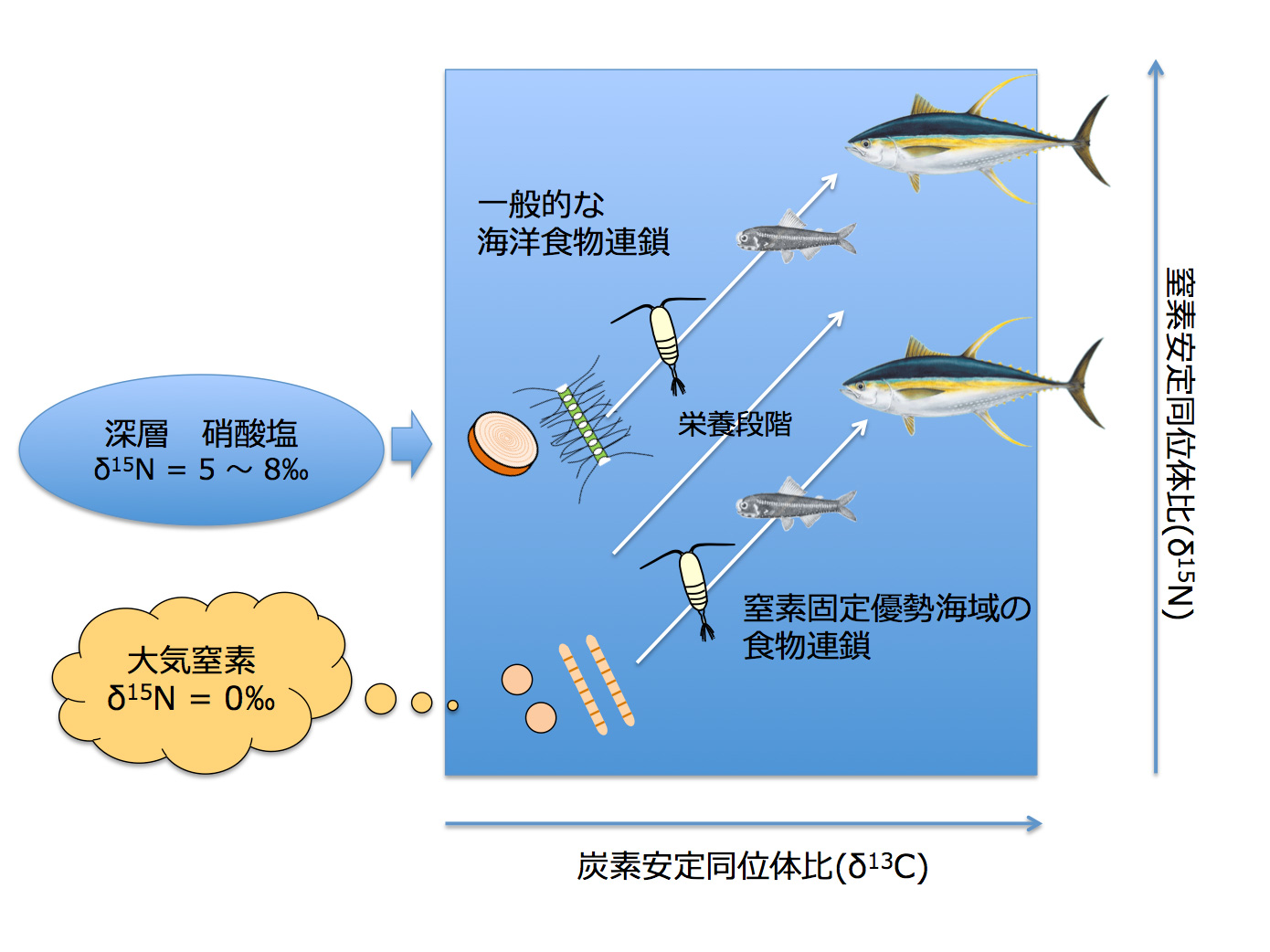
図4 今回発見された亜熱帯外洋域における窒素固定生物から高次捕食者に繋がる食物連鎖の概念図. (拡大画像↗)
亜熱帯外洋域表層は栄養塩濃度が極めて乏しいため、クロロフィル濃度(注1)が低く「海の砂漠」と比喩されています。この貧栄養海域において植物生産に特に足りないのが窒素栄養塩です。このような貧栄養海域の窒素供給プロセスとして近年窒素固定(注2)が注目を集めてきました。窒素固定とは窒素分子を還元してアンモニアを合成するプロセスで、特殊な微生物によって行われる生物学的プロセスです。窒素固定が貧栄養海域の窒素栄養塩として重要な役割を果たしていることがわかったのは、1990年代末とごく最近のことですが、それ以降、海洋窒素固定の研究は大いに進展し、全球的な物質循環を考える上で窒素固定生物の役割は極めて重要であることが明らかになってきました。一方、食物連鎖の観点から、窒素固定者の生産物(窒素)が高次栄養段階、すなわち水産資源生産に寄与しているのかという点にも注目が集まっていましたが、窒素固定生物の捕食者の特定は技術的に困難な点が多く、その研究は十分に進展していませんでした。
今回、東京大学大学院農学生命科学研究科、海洋研究開発機構、東京海洋大学らの研究グループは、中央太平洋において全長12,000kmの南北縦断観測線上の各観測点で、窒素固定速度および栄養塩濃度の実測に加えて、同じ測点から得られた植物プランクトン(基礎生産者:窒素固定者を含む)、植物プランクトンの捕食者である動物プランクトン、これを捕食する魚類マイクロネクトン(注3)の窒素・炭素安定同位体比(注4)を測定し、測点間で比較することにより、窒素固定者が亜熱帯外洋域表層食物連鎖に実際に寄与しているのかを検証しました。植物プランクトンの窒素安定同位体比は、利用した窒素源の同位体比を反映するため、窒素ガスを利用する窒素固定者が存在すると、その安定同位体比は、深層の硝酸塩(注5)を利用する他の植物プランクトンより軽くなることが知られています。また植物プランクトンを基点とする食物連鎖における窒素および炭素安定同位体比は、それぞれ栄養段階を経るごとに一定の割合で濃縮することが南極海や北太平洋亜寒帯海域での近年の研究から明らかにされてきました。これらの既往知見から、亜熱帯に窒素固定者を起点とする食物連鎖が存在しているのであれば、窒素および炭素安定同位体比のXYプロットにおいて、低い切片を持つ直線が窒素固定の活発な海域で検出されることが予想されます。
試料分析の結果、亜熱帯域の植物プランクトンの窒素安定同位体比は現場の窒素固定速度と有意な相関を示すことが明らかとなり(図1)、とくに低い海域が北緯15〜25度と南緯20〜30度と赤道を挟んで両半球に見られることが明らかとなりました(図2a)。これらの海域では、動物プランクトン、それらを捕食する魚類マイクロネクトンの安定同位体比も同様に他の海域より低く、食物連鎖を構成する群集全体が窒素固定者の生産に依存している可能性が示されました。一方亜熱帯域の植物プランクトンの炭素安定同位体比は、水温と正の相関を示しましたが、海域間の変動幅は僅かでした(図2b)。そこで各海域から得られた各食物連鎖構成要素の炭素・窒素安定同位体比の値を用いて、クラスター解析により各測点のグループ分けを行ったところ、亜熱帯海域の食物連鎖は、窒素固定者により支えられている海域(Group 2)、窒素固定がほとんどなく深層からの硝酸塩供給により支えられている海域(Group1)および両者の中間的な性質を示す海域(Group 3)に区分されることが明らかとなりました(図3)。これらの結果より、窒素固定の活発な海域(Group 2)では、その生産物が少なくとも魚類マイクロネクトンまで転送されていることが明らかとなりました。また、XYプロットにおける各回帰直線の傾き、すなわち各海域の食物連鎖過程における炭素窒素安定同位体比の濃縮率は、亜熱帯海域間で一定であるだけでなく、南極海や北太平洋亜寒帯で報告されている値とほぼ同じであることが明らかとなりました。このことは、外洋漂泳区では基礎生産者から上位栄養段階に至る食物連鎖において、炭素窒素の転送効率が、水温や群集構成種に関わらず一定であることを示しており、それぞれの食物連鎖に依存した高次捕食者の安定同位体比は、これらの回帰直線の延長線上にプロットされることを示唆しています。そこで、亜熱帯水産資源の代表種として、太平洋外洋に広く分布し、魚類マイクロネクトンを主な餌料としているキハダの炭素・窒素安定同位体比について、既往文献の調査を行ったところ、窒素固定が活発なことで知られるハワイ周辺のキハダが、本研究で示されたGroup 2の魚類マイクロネクトンの捕食者と推定される同位体比と一致する特徴を持っていることが明らかとなりました(図3)。このことはハワイのキハダは窒素固定者を起点とする食物連鎖上に位置していることを示唆しています(図4)。今回の研究結果は、一見生産性の低い亜熱帯外洋域が、なぜカツオ、マグロ類などを始めとする重要な水産資源の漁場となっているのか、そのメカニズムを考える上で、窒素固定者から始まる食物連鎖が重要な役割を果たしていることを示していると考えられます。
窒素固定が食糧生産に果たしている役割は、根粒菌とマメ科植物に代表されるように陸上生態系ではよく知られていますが、今回の研究結果は海洋においても陸上と同様に窒素固定者が食糧生産に深く関わっていることを示しています。亜熱帯海域の窒素固定者は種多様性が高く、温暖化や酸性化などの気候変動に対してそれぞれの種が異なる応答を示すことが予想されるため、今後は窒素固定者から始まる食物連鎖の詳細な構造を明らかにすることが、亜熱帯外洋域の魚類生産変動を理解する上で重要な研究課題となります。
発表雑誌
- 雑誌名
- :Global Ecology and Biogeography
- 論文タイトル
- :Stable isotopic evidence for differential contribution of diazotrophs to the epipelagic grazing food chain in the mid-Pacific Ocean
- 著者
- :Horii, Sachiko*; Takahashi, Kazutaka*; Shiozaki, Takuhei; Hashihama, Fuminori; Furuya, Ken
- DOI番号
- :10.1111/geb.12823
- 論文URL
- :https://onlinelibrary.wiley.com/doi/10.1111/geb.12823
問い合わせ先
准教授 高橋 一生(たかはし かずたか)
Tel:03-5841-5290
E-mail:akazutak<アット>mail.ecc.u-tokyo.ac.jp <アット>を@に変えてください。
用語解説
- 注1 クロロフィル濃度
- 植物光合成色素の一つ。すべての海洋植物プランクトンが共通してもつため、植物プランクトン現存量の指標として使われる。
- 注2 窒素固定
- 窒素ガスを還元してアンモニアを生成する過程。窒素ガスは化学的に非常に安定しているため、通常の植物は養分として利用できないが、一部の原核生物は窒素固定能をもつため、硝酸塩やアンモニウム塩などの窒素養分が枯渇した海域でも生育できる。
- 注3 魚類マイクロネクトン
- 外洋の中深層(200-1000m)に生息する小型魚類群(体長数センチ〜10センチ程度)。プランクトン(浮遊生物)とネクトン(遊泳生物)の中間の遊泳力をもち、一部の種は顕著な日周鉛直移動を示し、夜間表層で動物プランクトンを補食する。亜熱帯海域では大型魚類の主要な餌料生物のひとつ。
- 注4 安定同位体
- 同じ性質を持ちながら、重さ(質量数)の異なる元素。重さの違いからその反応速度に差が生まれるため、質量数の高い同位体(窒素の場合は15N)は生物の体内に残存しやすく、また捕食される度に一定の割合で濃縮することが知られている。この性質から標準物質(窒素の場合は大気中窒素ガス)との存在比が栄養段階の指標となる。
- 注5 硝酸塩
- 植物プランクトンが利用する主要な窒素源。海洋には莫大な量の硝酸塩が存在するが、表層付近は植物プランクトンに利用されるので、枯渇しやすい。このため、一般には下層から表層付近に供給される硝酸塩の多寡が海洋基礎生産を左右する。