| 2008/11/27 |
|
発表者: 中嶋 正敏(応用生命化学専攻・助教)
発表概要
植物ホルモン・ジベレリンに対する核内受容体GID1の構造を調べ、ジベレリン分子がいかなる結合様式に拠って受容体タンパク質に捕捉されるのかを解明した。
発表内容
ジベレリンは、種子の発芽・茎部の伸長・花器官の誘導分化・果実の成熟など種々の局面で植物の生長を制御する植物ホルモン(注1)の一種です。2005年に、複数の国内機関との共同研究により世界に先駆けてジベレリンに対する核内受容体GID1 (GIBBERELLIN INSENSITIVE DWARF1) がイネで発見されました(注2)。ジベレリンには130種を越える構造のよく似た類縁体が天然に存在しますが、それ自身がジベレリンとしての活性を有する「活性型ジベレリン」はGA(4)やGA(1), GA(3)など少数に限られます。試験管内実験系を用いてGID1に対する親和性を比較したところ、GA(1)やGA(3)に比べてGA(4)が最も高い親和性を示すこと、他方、いわゆる活性型ではないジベレリンはいずれもGID1に親和性を示さないことが判明しています。
本研究では大腸菌を用いて調製したイネGID1タンパク質と、GA(4)あるいはGA(3)との複合状態の結晶を調製し、X線結晶解析技術を用いてその構造を明らかにしました。この成果により、GID1タンパク質内のどのアミノ酸残基がジベレリンの認識に関わっているのか、なぜ活性型に属さないジベレリンはうまくGID1タンパク質と結合できないのか、などの疑問点に対して明快な説明が可能となりました。さらに私たちは、今回得られた情報に基づき点変異(注3)を導入した変異gid1タンパク質を調製して、活性型および非活性型のジベレリンとの親和性の変化を調べ、得られた構造情報の正しさを実証しています。
ジベレリンはいくつかの農産物の生産現場において活用される重要な植物生長調節剤です。その受容体タンパク質の構造が解き明かされたことは単に学術的な価値に留まらず、より効力の強いジベレリン様薬剤の開発や、ジベレリンと拮抗的な作用を示す薬剤の開発等においても極めて有益な情報を提供するものと期待されます。
添付資料
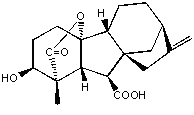
図 A :活性型ジベレリン GA(4) の化学構造
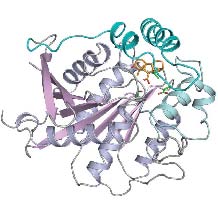
図B:ジベレリン受容体GID1と活性型GA(4)との複合体の結晶構造
※ GA(4)は中央右上に黄色で骨格構造が示されている
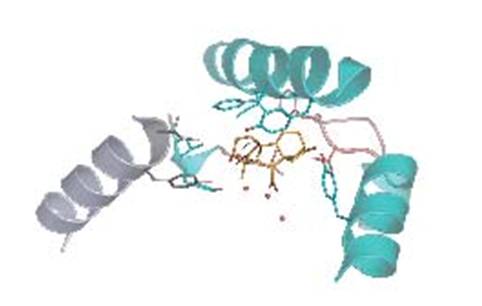
図C:GID1のジベレリン結合部位近傍と活性型GA(4)
発表雑誌
Asako Shimada, Miyako Ueguchi-Tanaka, Toru Nakatsu, Masatoshi Nakajima, Youichi Naoe, Hiroko Ohmiya, Hiroaki Kato and Makoto Matsuoka
Structural basis for gibberellin recognition by its receptor GID1
<共同研究者の所属情報>
| 松岡 信 | (名大生物機能開発利用研究センター教授) | |
| 上口(田中)美弥子 | (名大生物機能開発利用研究センター准教授) | |
| 島田麻子 | (名大生物機能開発利用研究センター) | |
| 大宮博子 | (名大生物機能開発利用研究センター) | |
| 加藤博章 | (京大院薬学研究科教授、理研播磨研究所) | |
| 中津 亨 | (京大院薬学研究科准教授、理研播磨研究所) | |
| 直江洋一 | (京大院薬学研究科) |
問い合わせ先
東京大学大学院農学生命科学研究科応用生命化学専攻
生物制御化学研究室 中嶋正敏
nkjm@pgr1.ch.a.u-tokyo.ac.jp
http://pgr1.ch.a.u-tokyo.ac.jp
用語解説
(注1)植物ホルモン
植物体内で生合成され、かつ極めて微量で植物の生長を制御する低分子化合物のことを「植物ホルモン」と呼ぶ。現在までに、上記ジベレリンの他に、オーキシン・サイトカイニン・エチレン・アブシジン酸・ブラシノステロイド・ジャスモン酸・ストリゴラクトンなどが知られている。また、これらに加えて近年ではペプチド性の生理活性物質もいくつか発見されている。
(注2)核内受容体GID1
イネ変異体プールの中から選抜された「背丈が非常に短く、かつ、ジベレリンを投与しても応答性を示さない変異体gid1」の原因遺伝子としてGID1 (GIBBERELLIN INSENSITIVE DWARF1)は特定された[Nature, 437: 693-698 (2005)]。GID1遺伝子の翻訳産物は活性型ジベレリンとの結合性を有しており、ジベレリンと受容体GID1が2者複合体を形成した場合に限り「ジベレリンからのシグナル伝達を常時抑制している制御因子」と新たに親和性を示すようになる。この3者複合体の形成によりシグナル伝達の抑止状態が解除され、その結果、ジベレリンに端を発するシグナル伝達が完了すると考えられている。
(注3)点変異
タンパク質を構成するアミノ酸残基の中で、その一部を本来とは異なる種のアミノ酸残基に変えて大腸菌などに作らせることができる。例えば1残基だけ(セリンからアラニンへなど)変換されたタンパク質を作った場合は「1点変異の導入」と表現される。分子内における特定のアミノ酸残基の機能を確認・解析する際などに有効な研究手法となっている。
〒113−8657 東京都文京区弥生1−1−1 www-admin@www.a.u-tokyo.ac.jp
Copyright © 1996- Graduate School of Agricultural and Life Sciences, The University of Tokyo