| 2009/7/10 |
|
発表者:
鈴木 浩之(応用生命化学専攻・大学院博士課程修了)
Park Seung-Hyun(応用生命化学専攻・博士研究員)
大久保 杏子(応用生命化学専攻・大学院修士課程修了)
北村 淳(応用生命化学専攻・大学院修士課程修了)
浅見 忠男(応用生命化学専攻・教授)
中嶋 正敏(応用生命化学専攻・助教)
発表概要
双子葉植物シロイヌナズナに存在する3種のジベレリン受容体のうち、2種が機能的に異常となった場合、その組み合わせに依って異なる形質が現れます。これは、シグナル伝達様式の複雑さを予想させる結果です。本研究により、「正常に機能する受容体の量的な欠如」と、「受容体の直下に位置するシグナル伝達因子への親和性の低さ、言い換えれば、相性の悪さ」が関係していることが明らかとなりました。
発表内容
ジベレリンは、種子の発芽・茎部の伸長・花器官の発達など種々の局面で植物の成長を制御している植物ホルモンです[注1・図A]。研究用植物として広く用いられるシロイヌナズナには、このジベレリンに対する受容体が全部で3種(GID1a, b, c)存在します[注2]。ジベレリンと複合体を形成した受容体はその時点から新たに「シグナルの伝達を常時抑制している制御因子DELLA」への親和性を生じ、その結果、ジベレリン-受容体-DELLA因子の3分子で構成する高次複合体を形成します。この高次複合体の形成により、DELLA因子の機能が弱まり、伝達抑制状態からの解除がジベレリンに端を発するシグナルの伝達開始に繋がっていると解釈されています[図B]。受容体間では機能的に重複が多く、いずれか1種に異常が生じても植物に異常な形質は現れませんが、3種のうちから2種同時に異常を来した場合(二重変異体2KO)には、特定の組み合わせでしかも限られた器官のみに異常形質が現れます。そこで、こうした機能的分担制御の仕組みを解明するため、受容体遺伝子にGUS酵素遺伝子を融合させ、受容体遺伝子のプロモーターでその発現を制御させる形質転換体を作出して、GUS遺伝子の発現状況およびGUS酵素活性の分布状況を調べました。
GID1aとGID1cが異常となった二重変異体(gid1a gid1c-2KO)では草丈のみ異常が現れ、花茎の伸長不良が生じます。そこで、この植物体で唯一正常に機能するGID1bに注目しました。GID1b-GUS遺伝子を発現する形質転換体の花茎では、遺伝子の発現は有意に認められたのに対してGUS酵素の活性は全く認められなかったことから、GID1bが安定して存在しないことが花茎で形質の現れた要因と判明しました。
別の二重変異体gid1a gid1b-2KO(GID1aとGID1bが異常となったもの)では花茎は正常ですが雄しべの発達不全を生じ、花粉が雌しべ柱頭まで届かないために深刻な不稔を招きます[図C]。この植物体で唯一正常に機能するGID1cに注目して、GID1c-GUS遺伝子を発現する形質転換体の雄しべにおけるGUS酵素活性を調べたところ、先のケースと異なり十分に活性が認められました。つまり、雄しべではGID1cが量的に欠乏するとは考えにくい結果が得られました。そこで、競合的酵母three-hybridと呼ばれるシステム[図D]を構築して、全部で5種存在するDELLA因子と受容体間の結合に関する親和性の強弱を調べました。その結果、雄しべにおける主要DELLA因子に対してGID1cが高い親和性を示す組み合わせが存在せず、比較的解離しやすい高次複合体の形成は主要DELLA因子の機能的回復につながるため、言い換えれば、シグナル伝達抑制状態が継続されることにより異常形質が生じたと考えられます[図E]。
ジベレリンはブドウやイチゴなど農産物の生長調節のために広く生産現場で利用されています。本研究は、受容体・DELLA因子がそれぞれ複数存在するような複雑な制御系であっても、各因子量と両因子間の親和性(「量」と「質」)の把握により制御機構の解釈が可能であることを示しており、その情報は特定部位に限定したシグナル制御技術の開発に道筋をつけるものです。
添付資料
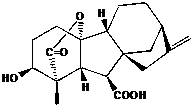
図A:代表的な活性型ジベレリンGA(4)の化学構造
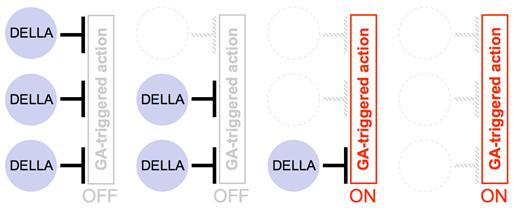
図B:ジベレリンのシグナル伝達の要点
受容体はジベレリンと結合することにより、新たにDELLA因子に対して親和性を示すように性状が変化する。ジベレリン-受容体-DELLA因子で構成される3分子複合体の形成により、DELLA因子のシグナル抑制機能が弱められる。ジベレリンに端を発したシグナルが最終的に伝達されるか否かは、その部位・器官に存在するDELLA因子の機能をジベレリン受容体が総量的にどの程度まで弱められるかに依存しており、「ジベレリン受容体に捕捉されていない状態のDELLA因子」の存在量をある閾値以下に減らすことができればシグナルが伝達され始める。

図C:gid1a gid1b-2KOの雄しべに現れた異常形質
変異体の雄しべ(右)は正常個体の雄しべ(左)と比較して伸長が不良で、雌しべの先端(柱頭)より下の側面に花粉が付く。この受粉効率の悪さは不稔を招く。
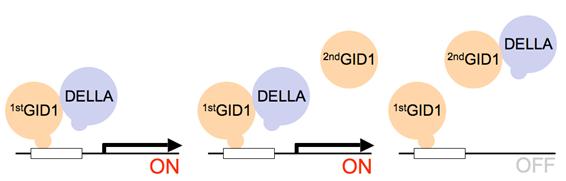
図D:競合的酵母three-hybrid系
ジベレリンの介在により受容体(1st)とDELLA因子が複合体を形成した場合(左図)、幾つかの遺伝子発現が開始されるように仕組まれており、それに伴って通常なら生育が制限される環境下でも上記複合体を形成した酵母のみ生育が可能となる。この環境に別種のジベレリン受容体(2nd)が共存する場合、受容体(1st)からDELLA因子を奪うほどの親和性を受容体(2nd)が持たなければそのまま生育可能な状況が継続される(中図)。しかし、DELLA因子に対する親和性が高いために、受容体(2nd)が受容体(1st)からDELLA因子を奪う状況が維持されると生育に必要な遺伝子の発現が途絶する(右図)。従って、酵母の生育状況を調べることにより3分子間の親和性を評価することが可能となる。
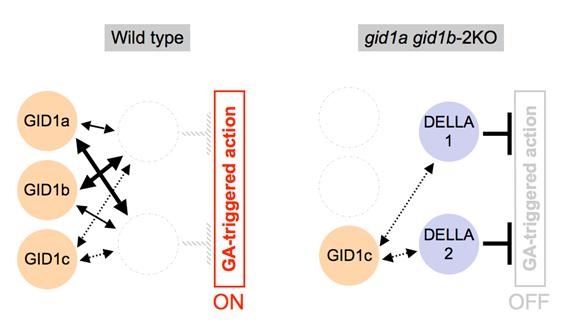
図E:gid1a gid1b-2KOの雄しべで異常形質が現れる仕組み
競合的酵母three-hybrid系を用いて評価した「受容体-DELLA因子間の親和性」をもとに、親和性の高低を線の太さで表現した。シロイヌナズナの雄しべに存在する主要DELLA因子にとってGID1cに高い親和性を示すものが含まれないことから、gid1a gid1b-2KO二重変異体においてのみジベレリンからのシグナルが十分に伝わらず、その結果、雄しべの伸長不良が生じたと考えられる。
発表雑誌
Differential expression and affinities of Arabidopsis gibberellin receptors can explain variation in phenotypes of multiple knock-out mutants.
Hiroyuki Suzuki, Seung-Hyun Park, Kyoko Okubo, Jun Kitamura, Miyako Ueguchi-Tanaka, Satoshi Iuchi, Etsuko Katoh, Masatomo Kobayashi, Isomaro Yamaguchi, Makoto Matsuoka, Tadao Asami and Masatoshi Nakajima
学外共同研究者の所属情報
上口(田中)美弥子 (名大生物機能開発利用研究センター・准教授)
井内 聖 (理化学研究所バイオリソースセンター・専任研究員)
加藤 悦子 ((独)農業生物資源研究所・主任研究員)
小林 正智 (理化学研究所バイオリソースセンター・主任研究員)
山口五十麿 (前橋工科大学・教授)
松岡 信 (名大生物機能開発利用研究センター・教授)
問い合わせ先
東京大学大学院農学生命科学研究科応用生命化学専攻
生物制御化学研究室 中嶋 正敏
nkjm[@]pgr1.ch.a.u-tokyo.ac.jp
http://pgr1.ch.a.u-tokyo.ac.jp
用語解説
[注1] 植物ホルモン
植物の体内で生合成され、微量で自身の成長・分化を制御する低分子化合物の総称。現在までに、ジベレリンの他、オーキシン・サイトカイニン・エチレン・アブシジン酸・ブラシノステロイド・ジャスモン酸・ストリゴラクトンなどが知られる。加えて近年、様々なペプチド性制御物質も植物の成長・分化に関与することが明らかになってきている。
[注2] ジベレリン受容体
イネ変異体プールから「背丈が非常に短く、かつ、投与されたジベレリンに対して応答性を示さない変異体gid1」が選抜され、その原因遺伝子GIBBERELLIN INSENSITIVE DWARF1(GID1)はジベレリンに対する受容体をコードしていた。膜貫通領域は持たず、可溶性のタンパク質である。
〒113−8657 東京都文京区弥生1−1−1 www-admin@www.a.u-tokyo.ac.jp
Copyright © 1996- Graduate School of Agricultural and Life Sciences, The University of Tokyo