| 2008/6/17 |
|
発表者: 有村慎一(生産・環境生物学専攻植物分子遺伝学研究室)
藤本優(同)
洞庭葉子(同)
角谷直紀(同)
中園幹生(同)
坂本亘(岡山大学資源生物科学研究所)
堤伸浩(生産・環境生物学専攻植物分子遺伝学研究室)
発表概要
植物細胞のミトコンドリアが分裂するために必要な新規因子 ELM1 を発見しました。 ELM1 は植物特異的であり、 DRP3A という真核生物共通のミトコンドリア分裂遂行因子を細胞質からミトコンドリアへ局在させる働きがあることがわかりました。この知見によって、植物細胞でのミトコンドリアの増殖や機能のさらなる解明につながります。
発表内容
植物細胞内でエネルギー生産、代謝、葉緑体のストレス緩和等の重要な役割を担うミトコンドリアは、細胞内で新たに作られることはなく既存のミトコンドリアが分裂することによって増殖していきます。植物ミトコンドリアの分裂・増殖機構の研究は、広く真核生物全体の細胞内エネルギー生産プラントの維持・増殖機構の解明につながります。
ミトコンドリアの分裂にはダイナミン様タンパク質という真核生物一般に広く保存されている因子がポリマー化して分裂面に巻きつき、ミトコンドリアをくびりきるように切断分裂させることが知られています。我々は以前、研究モデル植物であるシロイヌナズナを使って、ミトコンドリア分裂にかかわるダイナミン様タンパク質遺伝子 DRP3A を発見同定しました。その後さらに、 DRP3A 以外にも様々な因子がミトコンドリアの分裂に関与していると考え、ミトコンドリア分裂に不具合があると思われる複数の突然変異体シロイヌナズナを探索、発見してきました。今回は、その突然変異体の解析から新規因子 ELM1 を同定し、その役割を解明し報告しました。
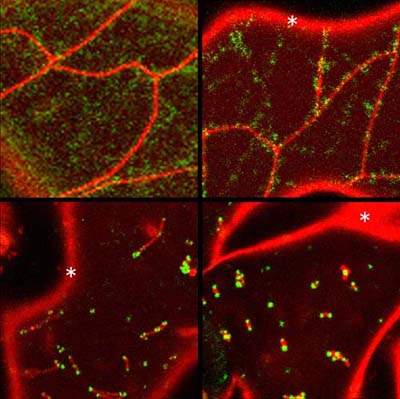
ミトコンドリアは無色透明であり顕微鏡下でもそのままでは判別しにくいため、 GFP (緑色蛍光タンパク質)をミトコンドリアへ局在させ、ミトコンドリアが緑の蛍光でラベルされた形質転換シロイヌナズナを作製しました。そしてこれに突然変異原処理をし、後代植物 19,000 個体の中から、ミトコンドリア分裂に不具合があると思われる(ミトコンドリアの数が減って長い)個体を発見し、その中の二系統をさらに解析しました。一つの系統は以前私たちが同定した DRP3A 遺伝子が機能不全になった drp3a 変異体でした。もう一つが elm1 ( el ongated m itochondria) 変異体です。分子生物学的な解析によって、 ELM1 タンパク質は DRP3A と結合することがわかり、また ELM1 はいつもミトコンドリアの膜上にいることがわかりました。興味深いことに elm1 突然変異体の中では、 DRP3A がミトコンドリアへ局在できず細胞質にとどまっていることが観察されました。ダイナミン様タンパク質は細胞質とミトコンドリア分裂面を行き来していることが知られていますので、 ELM1 が DRP3A の細胞質からミトコンドリア分裂面への局在に機能していることが推察されました。そこで、 elm1 突然変異体に ELM1 遺伝子を薬剤で誘導発現させることのできる形質転換植物を作製してこれを観察したところ、実際に ELM1 遺伝子の発現に応じて DRP3A が細胞質からミトコンドリアへ徐々に局在していき、 DRP3A が局在した箇所でミトコンドリアの分裂が起こることを証明することもできました(添付資料参照)。この形質転換植物の作製によって、ミトコンドリアの分裂とミトコンドリア形態を人為的にコントロールすることに成功したことにもなります。今後、これらの知見とこの形質転換植物体を使うことによって、植物ミトコンドリア分裂・形態の機能・生物学的意義などをさらに深く探ることができると考えています。
添付資料
発表雑誌
Arabidopsis ELONGATED MITOCHONDRIA1 Is Required for Localization of DYNAMIN-RELATED PROTEIN3A to Mitochondrial Fission Sites.
Shin-ichi Arimura, Masaru Fujimoto, Yoko Doniwa, Naoki Kadoya, Mikio Nakazono, Wataru Sakamoto, and Nobuhiro Tsutsumi
注意事項
特に無し。
問い合わせ先
堤伸浩 東京大学大学院農学生命科学研究科 生産・環境生物学専攻 植物分子遺伝学 教授
用語解説
〒113-8657 東京都文京区弥生1-1-1 www-admin@www.a.u-tokyo.ac.jp
Copyright © 1996- Graduate School of Agricultural and Life Sciences, The University of Tokyo