注目を集める希少放線菌の不思議に迫る研究
- 発表者
-
毛利 佳弘(東京大学大学院農学生命科学研究科応用生命工学専攻 博士課程(当時))
小西 健司(東京大学大学院農学生命科学研究科応用生命工学専攻 修士課程(当時))
藤田 梓(東京大学大学院農学生命科学研究科応用生命工学専攻 修士課程(当時))
手塚 武揚(東京大学大学院農学生命科学研究科応用生命工学専攻 助教)
大西 康夫(東京大学大学院農学生命科学研究科応用生命工学専攻 教授)
発表のポイント
◆アクチノプラネス属希少放線菌の特徴的な胞子嚢形成に関する遺伝子の発現制御機構に、はじめて分子生物学のメスが入りました。胞子を作る放線菌であれば必ずもっている転写因子BldDが、アクチノプラネス・ミズーリエンシスの胞子嚢形成の開始を制御していることが示され、BldD標的遺伝子の網羅的解析が行われました。一連の研究は大きな注目を集めています。
発表概要
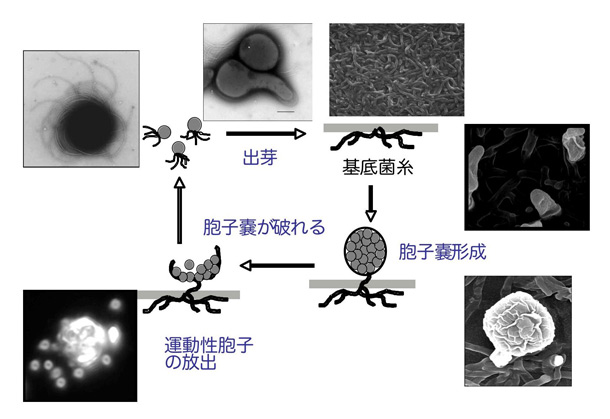
図1 希少放線菌アクチノプラネス・ミズーリエンシスの生活環
(拡大画像↗)

図2 今回の胞子嚢形成を制御する転写因子に関する研究については、希少放線菌の形態分化に関する最初の分子生物学的研究として“Commentary”で紹介されるとともに(文献6)、関連の電子顕微鏡写真がジャーナルの表紙を飾りました。
(拡大画像↗)
放線菌は抗生物質をはじめとする薬を作る微生物として知られていますが、細菌でありながらカビのように菌糸状に生育し胞子を形成するという点で、細胞分化や形態形成の仕組みの研究上、大変興味深い微生物です。これまで、ストレプトマイセス属放線菌を対象とした分子生物学的研究によって、胞子形成に必要な遺伝子が明らかにされ、その機能や発現制御機構などが詳細に研究されてきましたが、ストレプトマイセス属放線菌以外の放線菌に関しては、ほとんど研究されていませんでした。アクチノプラネス属放線菌は分離頻度の低い「希少放線菌」ですが、ストレプトマイセス属放線菌とは異なる生活環を有しています(図1)。アクチノプラネス属放線菌は、栄養増殖細胞である基底菌糸から、胞子嚢柄を介して、数百個の胞子を含む胞子嚢を形成します。湿潤した環境で胞子嚢は開裂し、胞子が放出されますが、この胞子は「べん毛」(注1)を持っており、高速で泳ぐことができます。走化性により、発芽に適した環境に辿り着いた運動性胞子(遊走子)は、運動を停止して発芽し、再び菌糸生育を始めます。アクチノプラネス属放線菌は、進化の中で、最も高度な形態分化能を獲得してきた細菌の一つと言えるでしょう。
我々の研究室では、アクチノプラネス属放線菌の形態分化の制御機構を研究することで、細胞の休眠や覚醒、形態分化の謎に迫ることができるのではないかと考え、約10年前にこの研究を開始しました。他のグループと共同でActinoplanes missouriensis(アクチノプラネス・ミズーリエンシス)の完全長ゲノム配列を決定するとともに、形質転換系の構築にも取り組みました(文献1、2)。また、胞子べん毛の観察も行いました(文献3)。昨年度、胞子べん毛の遺伝子およびその発現に関する研究をJournal of Bacteriology誌(注2)に報告したところ(文献4)、“Spotlight”として編集者に取り上げられ紹介されました(文献5)。今回の胞子嚢形成を制御する転写因子(注3)に関する研究については、希少放線菌の形態分化に関する最初の分子生物学的研究として“Commentary”で紹介されるとともに(文献6)、関連の電子顕微鏡写真がジャーナルの表紙を飾りました(図2)。このように、我々の希少放線菌の分子生物学的研究は、微生物学研究分野において、大きな注目を集めつつあります。
- Yamamura H, Ohnishi Y, Ishikawa J, Ichikawa N, Ikeda H, Sekine M, Harada T, Horinouchi S, Otoguro M, Tamura T, Suzuki K, Hoshino Y, Arisawa A, Nakagawa Y, Fujita N, Hayakawa M. Stand. Genomic. Sci.7(2): 294-303 (2012).
- Jang MS, Fujita A, Ikawa S, Hanawa K, Yamamura H, Tamura T, Hayakawa M, Tezuka T, Ohnishi Y. Plasmid 77: 32-38 (2015).
- Uchida K, Jang MS, Ohnishi Y, Horinouchi S, Hayakawa M, Fujita N, Aizawa S. Appl. Environ. Microbiol. 77(7): 2559-2562 (2011).
- Jang MS, Mouri Y, Uchida K, Aizawa S, Hayakawa M, Fujita N, Tezuka T, Ohnishi Y. J. Bacteriol.198(16): 2219-2227 (2016).
- Spotlight: Articles of Significant Interest Selected from This Issue by the Editors J. Bacteriol. 198(16): 2144(2016).
- Buttner MJ. J. Bacteriol. 199(12): e00070-17 (2017).
発表内容
転写因子BldDは、ストレプトマイセス属放線菌の形態分化のマスターレギュレーターの一つとして研究されてきました。BldDは胞子形成のための特殊な菌糸である気中菌糸の形成に必要な遺伝子群の転写を抑制しており、気中菌糸形成が起こるべき環境では、その抑制が解除されることで、気中菌糸形成の引き金が引かれます。胞子を形成する放線菌はBldDを持っていますが、ストレプトマイセス属放線菌以外では、その機能については、一種を除いて全く研究されていませんでした。
我々は希少放線菌アクチノプラネス・ミズーリエンシスにおいて、運動性胞子ではほとんど存在せず、発芽胞子に比較的大量に存在するタンパク質としてBldDを見出し、その機能解析を行いました。アクチノプラネス・ミズーリエンシスは、気中菌糸を形成せず、基底菌糸上に短い胞子嚢柄を介して、直接胞子嚢を形成しますが、この菌のBldDは不適切なタイミングでの胞子嚢形成の開始を抑制していることが、遺伝子破壊株を用いた解析から明らかになりました。また、細胞内でのBldD結合配列を網羅的に解析し、(1) BldDが染色体DNA上、少なくとも346箇所に結合していること、(2) その結合コンセンサス配列は19塩基からなる回文配列5' -nn(G/A)TnACn(C/G)n(G/C)nGTnA(C/T)nn-3'であること、(3) BldDの標的候補遺伝子には、転写因子をコードする遺伝子が多く含まれていること、を明らかにしました。また、標的候補遺伝子のうち27個について転写解析を行い、12個の遺伝子はBldDによって転写が抑制されていることを示しました。この中には、ストレプトマイセス属放線菌の形態分化に関与する遺伝子に相当するものが5つ含まれていました。一方、BldDによって制御される遺伝子群は、ストレプトマイセス属放線菌とアクチノプラネス・ミズーリエンシスでは、一部は共通であるものの、かなりの部分が異なっていることが示されました。
本研究成果は胞子形成を行う放線菌において、不適切なタンミングでの形態分化の開始を抑制するというBldDの機能の普遍性を示すものであると同時に、その標的遺伝子群は属ごとでかなりの相違があることを示した点で重要です。今後、BldD標的遺伝子を解析することで、アクチノプラネス・ミズーリエンシスの胞子嚢形成に関わる新しい遺伝子が見つかると期待されます。このようにして、放線菌に一般的な制御機構とアクチノプラネス・ミズーリエンシスに独特の制御機構を一つ一つ明らかにしていくことで、放線菌進化と微生物の生存戦略の謎に迫ることができると考えられます。また、そのような研究の中で、合成生物学における有用なツールや新しい機能性生物素材などの開発に資する成果が得られることが期待できます。
発表雑誌
- 雑誌名
- :Journal of Bacteriology, Volume 199, Number 12, e00840-16 (2017年3月27日オンライン版掲載)
- 論文タイトル
- :Regulation of sporangium formation by BldD in the rare actinomycete Actinoplanes missouriensis
- 著者
- :Yoshihiro Mouri, Kenji Konishi, Azusa Fujita, Takeaki Tezuka, Yasuo Ohnishi
- DOI番号
- :10.1128/JB.00840-16
- 論文URL
- :http://jb.asm.org/content/early/2017/03/21/JB.00840-16.abstract
問い合わせ先
- 東京大学大学院農学生命科学研究科応用生命工学専攻 醗酵学研究室
教授 大西康夫 - Tel:03-5841-5123
Email: ayasuo<アット>mail.ecc.u-tokyo.ac.jp <アット>を@に変えてください。
助教 手塚武揚
Tel: 03-5841-5126
Email: atezuka<アット>mail.ecc.u-tokyo.ac.jp <アット>を@に変えてください。
用語解説
- 注1 べん毛
- 細菌細胞から生えている毛のことで細菌の遊走のために使われる。ミドリムシや動物精子の運動器官である鞭毛では、鞭毛自身が構成タンパク質の収縮運動によって鞭打つように動くことで細胞に運動性を付与するのに対して、細菌のべん毛は付け根の部分がモーターとなっており、毛を回転させることで細胞に推進力を与える。つまり、細菌のべん毛は「鞭」ではないため、「鞭毛」ではなく「べん毛」と表記される。
- 注2 Journal of Bacteriology誌
- アメリカ微生物学会(American Society for Microbiology)が出版するジャーナルで1916年創刊と100年以上の歴史がある。基礎微生物学研究分野の最も重要なジャーナルの1つとして知られている。
- 注3 転写因子
- DNAに書き込まれた遺伝子情報は、RNAポリメラーゼという酵素によって、 mRNAとして読み取られるが、このステップは転写と呼ばれる。遺伝子発現の制御は転写段階で行われることが多く、転写のオン・オフの切り換えに関わるタンパク質を転写因子と呼ぶ。