新しい酵素サブファミリーを提唱:ポリエンを合成するII型ポリケタイド合成酵素
- 発表者
-
ト タンヤオ(東京大学大学院農学生命科学研究科 応用生命工学専攻 博士課程2年)
勝山 陽平(東京大学大学院農学生命科学研究科 応用生命工学専攻 准教授)
新家 一男(産業技術総合研究所 上級主任研究員)
大西 康夫(東京大学大学院農学生命科学研究科 応用生命工学専攻 教授)
発表のポイント
◆微生物が作るイシガミドという天然化合物の生合成に関わる酵素がポリエン骨格の合成反応を触媒していることを、試験管内酵素反応によって初めて証明しました。
◆本酵素はII型ポリケタイド合成酵素(II型PKS)ファミリーに属しますが、これまでII型PKSは芳香族化合物の生合成を担うと考えられてきたため、本酵素がより還元度の高い化合物であるポリエンを合成できることを証明した意義は大きく、「高還元型II型PKS」という新しいサブファミリーを提唱することができました。
◆本研究により、高還元型II型PKSが有するユニークな触媒メカニズムの解明の端緒が開かれました。
発表概要
II型ポリケタイド(注1)合成酵素(II型PKS)は、これまで芳香族化合物の生合成を担うと考えられてきました。一方、東京大学醗酵学研究室および産業技術総合研究所が以前行った共同研究により、比較的還元度の高いポリエン骨格を持つポリケタイド(イシガミドと命名)の生合成において、ポリエン骨格(注2)の合成を担うと考えられるII型PKS(IgaPKS)が放線菌(注3)から発見されました1。今回、同共同研究グループはIgaPKSの構成要素である5つのタンパク質を調製し、これらを用いることでポリエン骨格生合成経路を試験管内で再構築することに成功しました。一連の結果は、「II型PKSは還元度の低い芳香族化合物を生合成する」というこれまでの常識を覆すものであり、II型PKSの中には、高還元度のポリエンの生合成を担うものがあることが明確に示されました。さらに、分子系統解析(注4)により、IgaPKSおよびポリエン骨格の生合成を担うと予想される複数のII型PKSが1つのサブファミリーを形成していることが示され、「高還元型II型PKSサブファミリー」が提唱されました。本研究により、高還元型II型PKSが有するユニークな触媒メカニズムの解明の端緒が開かれました。
参考文献
1. D. Du, Y. Katsuyama, H. Onaka, M. Fujie, N. Satoh, K. Shin-Ya, Y. Ohnishi. Production of a Novel Amide-Containing Polyene by Activating a Cryptic Biosynthetic Gene Cluster in Streptomyces sp. MSC090213JE08. Chembiochem 17: 1464–1471, 2016.
発表内容
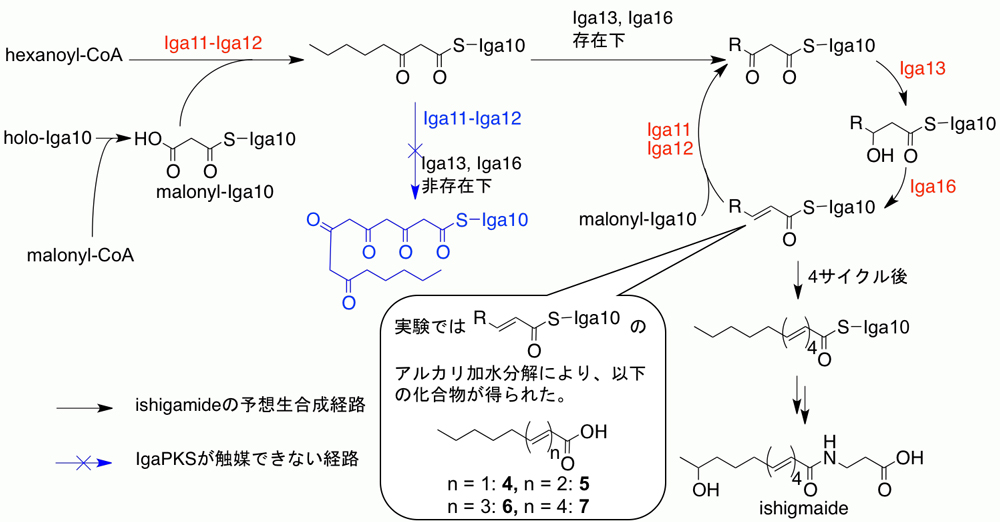
図1 イシガミドの生合成経路 (拡大画像↗)
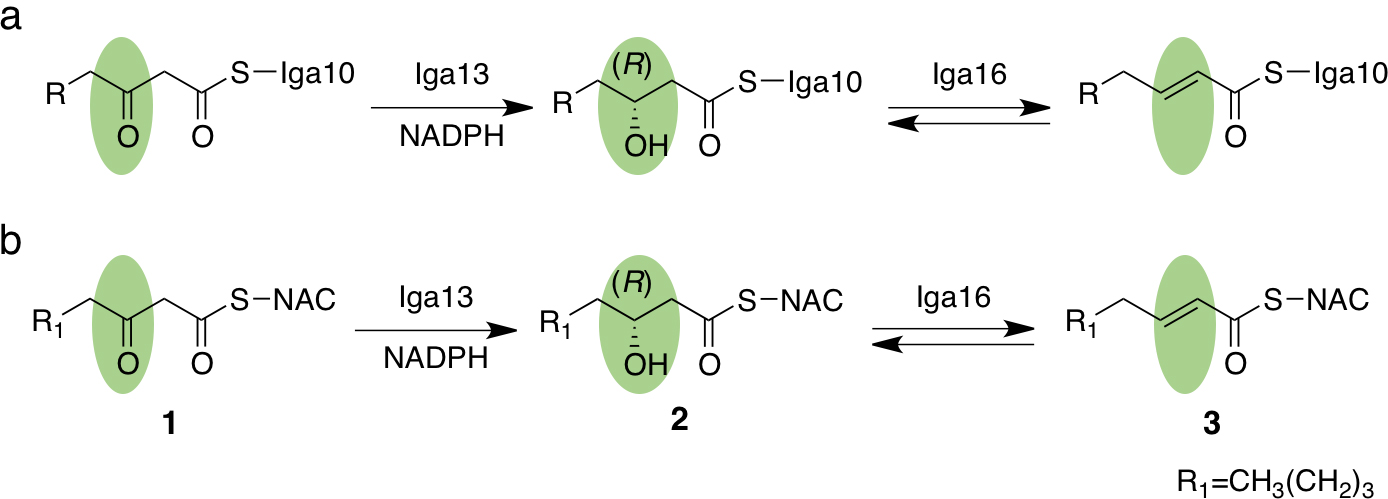
図2 Iga13とIga16が触媒する反応 a, 生体内におけるIga13とIga16が触媒する反応。b, 今回、基質アナログを用いて証明されたIga13とIga16が触媒する反応。 (拡大画像↗)
ポリケタイド合成酵素(PKS)が作り上げるポリケタイドは医薬品をはじめとする有用化合物の資源として重要であり、PKSに関する研究は古くから世界中で精力的に行われてきました。PKSはその構造および反応様式により、I型、II型、III型に分類されています。I型PKSは複数の機能ドメインを持つ1つの巨大タンパク質である一方、II型PKSは異なる機能を持つ複数のタンパク質から構成されます。I型PKSはさらにモジュラー型と反復型に分類されます。反復型I型PKSはその生産物の還元度合いにより、さらに高還元型、部分還元型、非還元型に分類できます。一方、II型PKSにおいて、このような細分類はなされていませんでした。典型的なII型PKSでは、ケト合成酵素(ketosynthase, KS)と鎖長決定因子(chain length factor, CLF) からなる複合体(KS-CLF)が触媒する縮合反応によって、ポリケタイド鎖が伸長し、ポリケトメチレン鎖が合成されます。その後、このポリケトメチレン鎖はさまざまな修飾酵素による修飾を受け、比較的還元度の低い芳香族化合物となります。このため、強いて言うなら、II型PKSは、反復型I型PKSサブファミリーでいうところの部分還元型あるいは非還元型に相当すると考えられてきました。
しかしながら、ここ数年の間に、芳香族化合物よりも還元度の高いポリエンの生合成を触媒すると予想されるII型PKSがいくつか報告されました。東京大学醗酵学研究室および産業技術総合研究所が行った共同研究によっても、ポリエン骨格を持つ放線菌ポリケタイドであるイシガミドの生合成において、ポリエン骨格の合成を担うと考えられるII型PKS、IgaPKSが発見されました。一連の報告は、II型PKSの中にも「高還元型」と分類すべきものがあることを示すものです。興味深いことに、これらのII型PKSは、分子系統解析によって、従来の典型的なPKSとは進化的に異なったグループに属することも明らかになり、「高還元型II型PKS」の存在がますます現実味を帯びてきました。しかしながら、これらの「高還元型II型PKS」はいずれも、その触媒反応を試験管内で証明するには至っていなかったため、「高還元型II型PKS」を新しい酵素サブファミリーとするには不十分な状況でした。そこで、今回の研究では、IgaPKSの触媒反応を試験管内で明らかにすることを目指しました。IgaPKSはポリエンを合成するため、その触媒サイクルにおいて、鎖伸長と還元、脱水の修飾反応を交互に繰り返すと予想されます(図1)。
IgaPKSはIga10(アシル基運搬タンパク質、acyl carrier protein, ACP)、Iga11(KS)、Iga12(CLF)、Iga13(ケト還元酵素、ketoreductase, KR)、Iga16(脱水酵素、dehydratase, DH)の5つのタンパク質から構成されると予想されました。各タンパク質の機能予測から、イシガミドの生合成経路が次のように推定されました。まず、Iga11-Iga12が開始基質であるヘキサノイルCoAとIga10に結合したマロニル基のクライゼン縮合を触媒します。次にIga13が触媒するケトンの還元反応、Iga16が触媒する脱水反応により、二重結合が形成されます。Iga11-Iga12、Iga13、Iga16による縮合、還元、脱水反応からなる触媒サイクルが繰り返されることで、ポリエン骨格が伸長し、イシガミドの不飽和脂肪酸部位が生合成されます。その後、イシガミド生合成遺伝子クラスター中にコードされた酸化酵素によって不飽和脂肪酸に水酸基が導入され、イシガミド生産菌が有する未知の酵素により不飽和脂肪酸部位とβ-アラニンの縮合が触媒され、イシガミドが生合成されます(図1)。
推定した生合成経路を検証するために、まずIga13とIga16の組換えタンパク質を調製し、機能解析を行いました。これらの反応性を調べるために、Iga10に結合した反応中間体を模倣したN-アセチルシステアミン(N-acetylcysteamine, NAC)体(図2、化合物1、2、3)を化学合成し、試験管内反応によりIga13とIga16が触媒する反応を調べました。その結果、Iga13はNADPHを補酵素とし、3-オキソオクタノイル-NAC(3-oxooctanoyl-NAC、化合物1)を(3R)-ヒドロキシオクタノイル-NAC((3R)-hydroxyoctanoyl-NAC、化合物2)に還元する活性を示しました。また、Iga16は2とトランス-2-オクテノイル-NAC(trans-2-octenoyl-NAC、化合物3)間の可逆反応を触媒しました。以上の結果から、Iga13とIga16は厳密な立体選択性(注5)を示し、Iga10に結合している反応中間体のβ-ケト基を立体特異的にR体の水酸基、E体のオレフィンへと変換することが示されました。
次に、Iga12とIga11の活性を調べるために、大腸菌を用いてこれらの組換えタンパク質の取得を試みました。これらのタンパク質は単独で得ることはできず、Iga11-Iga12複合体としてのみ、調製可能でした。Iga11-Iga12複合体の活性を調べるため、Iga11-Iga12複合体の組換えタンパク質をIga10マロニルCoA(malonyl-CoA)とヘキサノイルCoA(hexanoyl-CoA)とともに反応させました。その結果、3-オキソオクタノイル-Iga10(3-oxooctanoyl-Iga10)のみが生成しました。このことから、Iga11-Iga12複合体は単独では一回の縮合反応は触媒できるが、それ以上の縮合反応を行うことができないことがわかりました。
次に、IgaPKS全体の触媒反応を調べるために、Iga11-Iga12複合体、Iga10、Iga13、Iga16をマロニルCoAとヘキサノイルCoA、NADPHとともに反応させました。生成物はIga10に結合した状態で存在すると考えられたため、反応液をアルカリ処理し、Iga10に付いているポリケタイド鎖を加水分解により切り離しました。得られた化合物を分析したところ、4つの不飽和カルボン酸(4-7、図1)の生成が観察されました。この結果から、Iga11-Iga12、Iga13、Iga16がそれぞれ触媒する縮合、還元、脱水の反応サイクルによりポリエン骨格が伸長していることが明らかになりました。
本研究により、IgaPKSがポリエン骨格の生合成を担うことが証明されました。これにより、IgaPKSと類似したII型PKS群を「高還元型II型PKSサブファミリー」と提唱することができました。これは、「II型PKSは還元度の低い芳香族化合物を生合成する」というこれまでの常識を覆すものであり、その意義は大変大きいといえます。また、本研究により、高還元型II型PKSが有するユニークな触媒メカニズムの解明の端緒が開かれました。今後、IgaPKSの構造解析などを通して、高還元型II型PKSの詳細な触媒機構が明らかになることが期待されます。
本研究は、日本学術振興会の科学研究費補助金(JP25108706, JP17H05432, JP17J09439)、および A3 Foresight Programの支援を受けて行われました。
発表雑誌
- 雑誌名
- :Angewandte Chemie International Edition (2018年1月17日オンライン公開)
- 論文タイトル
- :Reconstitution of a Type II Polyketide Synthase Catalyzing Polyene Formation
- 著者
- :Danyao Du, Yohei Katsuyama, Kazuo Shin-ya, Yasuo Ohnishi.
- DOI番号
- :10.1002/anie.201709636
- 論文URL
- :http://onlinelibrary.wiley.com/doi/10.1002/anie.201709636/full
問い合わせ先
教授 大西 康夫 (おおにし やすお)
Tel:03-5841-5123
Fax:03-5841-8021
准教授 勝山 陽平 (かつやま ようへい)
Tel: 03-5841-5124
Fax: 03-5841-8021
研究室URL:http://park.itc.u-tokyo.ac.jp/hakko/
用語解説
- 注1 ポリケタイド
- 酢酸ユニットを基本単位とした多様な構造を示す化合物の総称。植物や微生物などの二次代謝産物として生合成されており、多様な生理活性を示すため、医薬品資源として重要である。
- 注2 ポリエン
- 少なくとも3つの二重結合と単結合が交互に並んだ炭化水素鎖構造をもつ化合物の総称。
- 注3 放線菌
- 真正細菌の一群で、菌糸を放射状に伸長させて増殖することから名付けられた。このような形態学的な特徴に加え、多くの二次代謝産物を作り出す能力を持つことでも知られ、クスリを作る微生物として産業上、有用である。放線菌が生産するさまざまな抗生物質は、医療現場で欠かせないものになっている。
- 注4 分子系統解析
- 遺伝子の塩基配列あるいはタンパク質のアミノ酸配列の比較から、生物や遺伝子が進化してきた道筋(系統)を調べる手法。
- 注5 立体選択性
- ある化学反応の生成物として複数の立体異性体が考えられる場合に、ある特定の立体異性体が優先的に得られる反応の性質。