2018/03/27
麹菌において融合した細胞の可視化に成功
-株どうしの細胞融合の相性を決める不和合性を麹菌で初めて発見-
- 発表者
-
岡部 知弥(東京大学大学院農学生命科学研究科応用生命工学専攻 修士課程:当時)
片山 琢也(東京大学大学院農学生命科学研究科応用生命工学専攻 特任助教)
莫 濤寧(東京大学大学院農学生命科学研究科応用生命工学専攻 修士課程)
森 法子(東京大学大学院農学生命科学研究科応用生命工学専攻 修士課程)
金 鋒杰(南京林業大学 教授)
藤井 郁雄(大阪府立大学理学系研究科 教授)
岩下 和裕(独立行政法人 酒類総合研究所 主任研究員)
北本 勝ひこ(日本薬科大学 特任教授)
丸山 潤一(東京大学大学院農学生命科学研究科応用生命工学専攻 特任准教授)
発表のポイント
◆麹菌において融合した細胞を可視化する方法を確立し、多くの麹菌実用株が細胞融合を行う能力をもつことを明らかにしました。
◆株の組み合わせによって融合細胞が存在できない不和合性と呼ばれる現象があることを、麹菌で初めて発見しました。
◆細胞融合は有性生殖に必要な過程であり、不和合性のメカニズムの解明が交配育種による優良な性質をもつ麹菌株の開発につながると期待されます。
発表概要
麹菌は、我が国の醸造産業だけでなく、酵素や異種タンパク質などの物質生産に利用されている産業的に重要な微生物です。麹菌では有性世代が発見されておらず、異なる株を掛け合わせて優れた能力をもつ株を育種する交配が困難であることが長年の課題となっています。また、交配育種での有性生殖においては、異なる株どうしで細胞を融合させる必要があります。麹菌の細胞融合に関する研究は、東京大学応用微生物研究所(当時)の坂口謹一郎 東京大学名誉教授らによる1950年代の報告があるのみで、以降、長い間取り組まれることはありませんでした。
今回、東京大学大学院農学生命科学研究科の丸山潤一特任准教授らのグループは、麹菌で融合した細胞を可視化する方法を確立しました。この方法を用い、多くの麹菌実用株が細胞融合を行う能力をもつことを明らかにしました。また、麹菌の異なる株を組み合わせることで、融合細胞が存在することができない不和合性と呼ばれる現象を初めて発見しました。今後は、有性生殖において株の組み合わせを決める障害となる不和合性のメカニズムを解明することにより、麹菌の効率的な交配育種技術が開発されることが期待されます。
発表内容
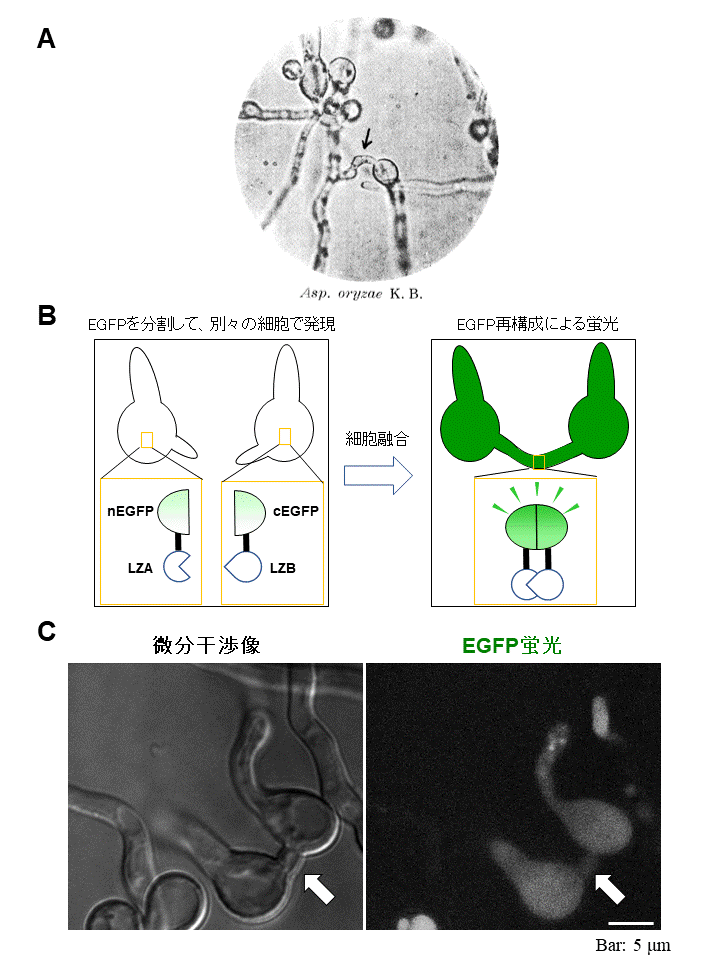
図1 BiFC法による融合細胞の可視化
(A) 1956年の論文に掲載された麹菌の融合した細胞の写真(Ishitani and Sakaguchi (1956) Journal of General and Applied Microbiology, Vol. 2, 345-400)。
(B) 融合細胞の可視化の原理。お互いに結合するタンパク質LZAとLZBに、EGFP蛍光タンパク質を2つに分割したnEGFPとcEGFPをそれぞれ融合させ、別々の細胞で発現させる。融合した細胞においてLZAとLZBが結合することより、EGFPタンパク質が再構成されて蛍光が観察される。
(C) 蛍光で可視化された融合細胞(矢印)。 (拡大画像↗)
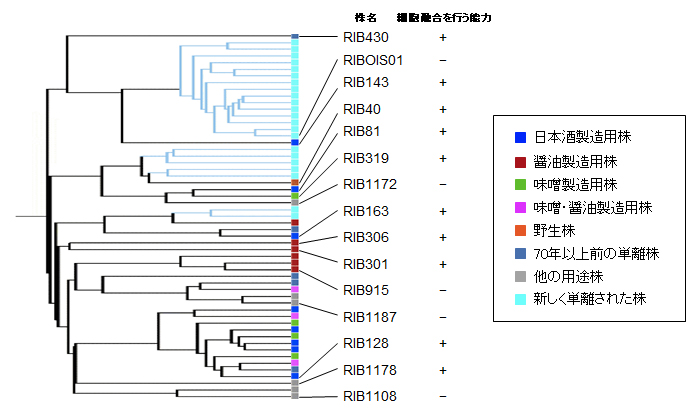
図2 麹菌株の系統樹と選んだ株の細胞融合能力の有無 (拡大画像↗)
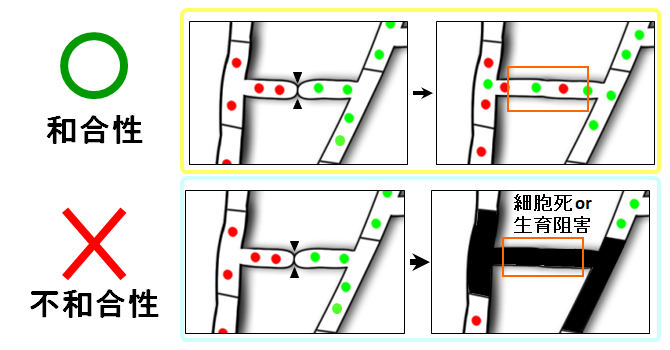 図3 糸状菌における和合性と不和合性
図3 糸状菌における和合性と不和合性
遺伝的に適合な株の細胞どうしが融合すると、融合細胞は安定に存在することができる(和合性)。不適合な株どうしが融合すると、細胞死または著しい生育阻害が誘導される(不和合性)。 (拡大画像↗)
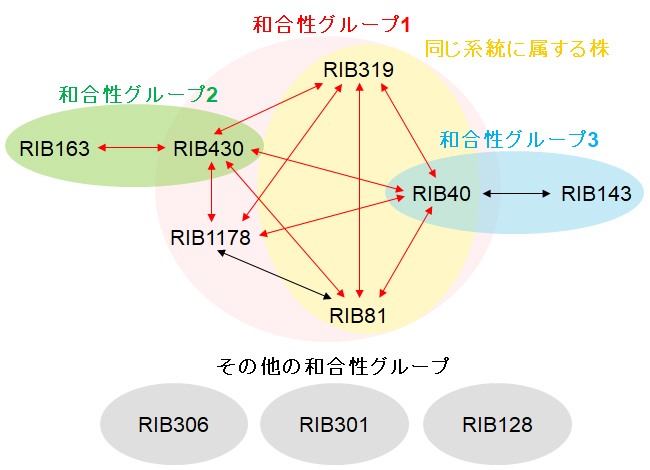
図4 麹菌株の和合性グループ
異なる株どうしの和合性・不和合性の組み合わせの判定結果をもとに、和合性グループに分類した。図2の系統図で同じ系統に属する株(RIB40, RIB81, RIB319)は、同じ和合性グループに属する。 (拡大画像↗)
麹菌は日本酒・醤油・味噌などの醸造食品の製造はもとより、酵素や異種タンパク質の生産に利用されている産業的に重要な糸状菌(カビ)です。麹菌では有性世代が発見されていないため、異なる株を掛け合わせてより優れた性質をもつ株を育種する交配を行うのは困難です。私たちは以前、麹菌には動物の雄雌の性別のような2つの接合型株が存在することを明らかにして、麹菌が有性生殖を行うことができる可能性を示しました。
有性生殖を行うには、それぞれの株の細胞どうしが融合することが必要です。麹菌の細胞融合については、1950年代に東京大学応用微生物研究所(当時)の坂口謹一郎 東京大学名誉教授、石谷(武市)千代子 博士らによる研究が行われていました。そのなかで、麹菌の細胞どうしが融合した写真が残されていますが(図1A)、麹菌の細胞融合に関する研究はそれ以降途絶え、長らく行われてきませんでした。最近、私たちは麹菌が細胞融合を行う能力をもつことを、約60年ぶりに再発見しました。しかし、その頻度は低く、融合した細胞の形態を観察することが困難でした。
そこで本研究では、蛍光タンパク質再構成法(Bi-molecular fluorescence complementation、以下BiFC法)(注1)を利用して、麹菌で融合した細胞を可視化して観察する方法を確立しました。このなかで、お互いに結合することができる2つのタンパク質に、緑色蛍光タンパク質EGFPを分割したものをそれぞれ融合して、別々の細胞に発現させました(図1B)。その結果、融合した細胞だけが蛍光を発し、その形態を観察することができました(図1C)。特に、分生子と呼ばれる無性胞子どうしが融合する形態が多く観察されました。これは、約60年ぶりに麹菌が細胞融合を行う際の形態を解明したものとして、特筆に値します。
麹菌は、日本酒・醤油・味噌などの用途によって様々な性質をもつ多くの実用株が使用されています。そこで、酒類総合研究所で作成された麹菌株の系統樹において、同じ系統もしくは異なる系統から14株の実用株および1株の野生株を選んで調べたところ、ほとんどの株が細胞融合を行う能力をもつことがわかりました(図2)。さらに、異なる株どうしを組み合わせて調べたところ、一部は融合しましたが、大部分の株の組み合わせでは細胞融合は観察されませんでした。
糸状菌では、遺伝的に不適合な株どうしで細胞が融合した場合、細胞死または生育の著しい低下が起こる不和合性という現象が知られています(図3)。上記の実験結果において、大部分の株の組み合わせで細胞融合が観察されなかったことから、麹菌においても不和合性の現象が存在する可能性が考えられました。そこで、プロトプラスト融合法(注2)によって異なる株の細胞どうしを強制的に融合させたところ、大部分の組み合わせで融合した細胞は観察されませんでした。この結果から、麹菌において株の組み合わせによって不和合性が存在することを初めて明らかにしました。そして、和合性・不和合性の組み合わせを検討することで、麹菌株を複数の和合性グループに分類することができました(図4)。そのうち1つの和合性グループでは、主に系統が近い株どうしで構成され、系統が離れた株は不和合性となる傾向があることがわかりました。
本研究では、融合細胞を可視化する方法を確立することによって、麹菌実用株の多くが細胞融合を行う能力をもつことを明らかにしました。また、麹菌における不和合性を初めて発見し、麹菌実用株を複数の和合性グループに分類することに成功しました。有性生殖には異なる株どうしの細胞融合が必要であることから、和合性グループの分類は組み合わせる麹菌株の選択をするために重要な情報となります。さらに今後、麹菌における不和合性を制御するメカニズムを解明することで不和合性を解消し、将来、効率的な交配育種技術が開発されることが期待されます。
発表雑誌
- 雑誌名
- :「Scientific Reports」
- 論文タイトル
- :BiFC-based visualisation system reveals cell fusion morphology and heterokaryon incompatibility in the filamentous fungus Aspergillus oryzae
- 著者
- :Tomoya Okabe, Takuya Katayama, Taoning Mo, Noriko Mori, Feng Jie Jin, Ikuo Fujii, Kazuhiro Iwashita, Katsuhiko Kitamoto, Jun-ichi Maruyama
- DOI番号
- :10.1038/s41598-018-21323-y
- 論文URL
- :https://www.nature.com/articles/s41598-018-21323-y
問い合わせ先
特任准教授 丸山 潤一(まるやま じゅんいち)
Tel:03-5841-5164
Fax:03-5841-8184
E-mail:amarujun<アット>mail.ecc.u-tokyo.ac.jp <アット>を@に変えてください。
用語解説
- 注1 蛍光タンパク質再構成法 (Bi-molecular fluorescence complementation、BiFC)
- 細胞内で2つのタンパク質間の結合を検出するために利用されます。結合を調べたい2つのタンパク質に、分割した蛍光タンパク質をそれぞれ融合して細胞に発現させると、それらのタンパク質が相互作用した場合にのみ分割された蛍光タンパク質が正常な構造をとって蛍光が観察されます。
- 注2 プロトプラスト融合法
- 真核微生物や植物の細胞壁を溶解して得られる細胞をプロトプラストと呼び、プロトプラスト懸濁液にポリエチレングリコール溶液を加えることで、細胞融合能に関わらずプロトプラストを融合させることのできる方法。