カイコ培養細胞を用いた人工ping-pong piRNA産生系の確立
- 発表者
-
庄司 佳祐(東京大学大学院農学生命科学研究科 生産・環境生物学専攻 博士課程3年;日本学術振興会特別研究員DC2)
鈴木 穣(東京大学大学院新領域創成科学研究科 メディカル情報生命専攻 教授)
菅野 純夫(東京大学大学院新領域創成科学研究科 メディカル情報生命専攻 教授)
嶋田 透(東京大学大学院農学生命科学研究科 生産・環境生物学専攻 教授)
勝間 進(東京大学大学院農学生命科学研究科 生産・環境生物学専攻 准教授)
発表のポイント
◆piRNAと呼ばれる小さなRNAを人工的に産生させる実験系を確立しました。
◆ping-pongサイクルと呼ばれるpiRNA産生経路の存在を実験的に示しました。
◆本研究で開発した人工piRNA産生系は、piRNAの配列に隠された特徴を見出すツールになることが期待されます。
発表概要
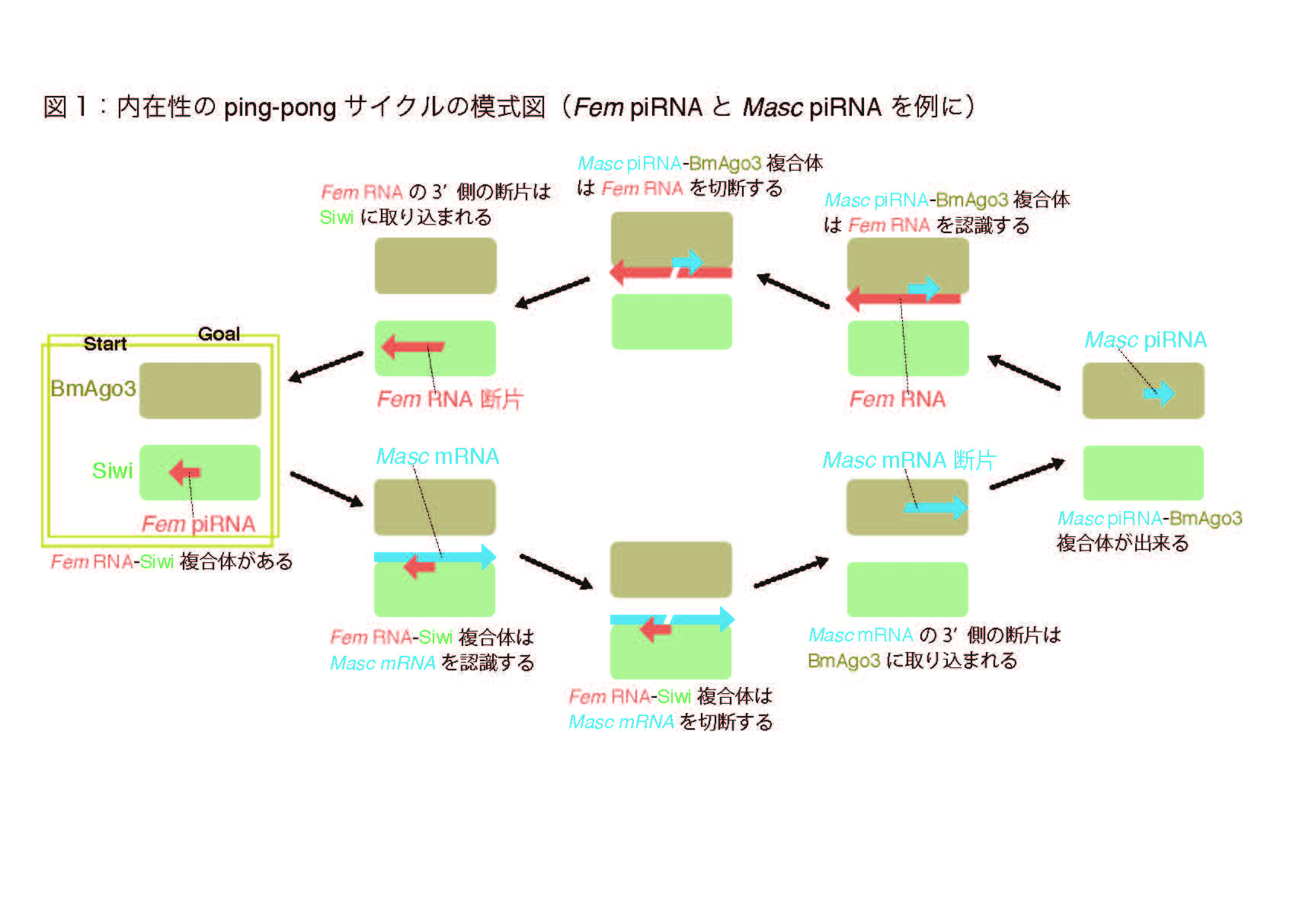
動物のゲノム中にはトランスポゾンと呼ばれる利己的な転移因子が潜んでいます。トランスポゾンは普段は抑制されていますが、その抑制が生殖細胞で解除されると生殖巣の発達が不全になってしまいます。このトランスポゾンの抑制の中核を担うのが、PIWI-interacting RNA(piRNA)と呼ばれる短いRNAです。これまでの研究から、piRNAの産生にはprimary pathwayとping-pong pathwayという2種類の経路が存在することが示唆されていました。ping-pong pathwayでは、piRNA-PIWI複合体によって切断された断片が新たなpiRNAの前駆体となり、新たなpiRNAとPIWI複合体による切断を受けたRNA が、更に新たなpiRNA(最初のものと同じ配列)の前駆体になるとされています(図1 )。しかし、この経路では同じ配列のペアが作られ続けるため、新しく作られたpiRNAを区別することは困難です。そのため、生殖細胞内で本当にping-pongサイクルが実在しているのかは実験的にしっかり示されてきませんでした。
今回、東京大学大学院農学生命科学研究科の勝間進准教授のグループは、人工的にping-pong piRNAを産生する実験系を構築することによって、ping-pongサイクルが実在することを示しました。さらに、この人工ping-pong piRNA産生系を用いることで、piRNAとターゲット配列がどの程度一致していれば認識できるのかを明らかにしました。本実験系は、piRNAに隠された配列上の特徴を見つける有用なツールであると考えられます。
発表内容
動物のゲノム中にはトランスポゾンと呼ばれる利己的な転移因子が潜んでいます。トランスポゾンは普段は抑制されていますが、その抑制が解除されると転移によって重要な遺伝子を破壊してしまうことがあります。そのため、遺伝情報を次世代へと伝達する生殖細胞では、トランスポゾンは適切に抑制しておく必要があります。この抑制の中核を担うのが、piRNAと呼ばれる30塩基に満たない短いRNAです。piRNAはPIWIタンパク質と結合して、piRNAと相補的な配列を持つmRNAを切断することでトランスポゾンなどの抑制を行います。そのため、piRNAの大部分はトランスポゾンmRNAと対応した配列を持っています。
piRNAの産生経路には、primary pathwayとping-pong pathwayという2種類の経路があると考えられています。バイオインフォマティクスによる配列解析から、ping-pong pathwayでは、piRNA-PIWI複合体によって切断されたRNA断片が新たなpiRNAの前駆体となり、そのpiRNAとPIWI複合体によるRNAの切断片が新たなpiRNA(最初のものと同じ配列)の前駆体になると考えられています(図1 )。このように、ある特定のpiRNAペアが増幅して行くプロセスは、卓球で球が行ったり来たりする様子に例えて、ping-pongサイクルと名付けられています。しかし、最初のpiRNAとping-pongサイクルの結果作り出されるpiRNAが全く同じ配列であること、また前駆体となるトランスポゾン配列の種類が非常に多様であることから、ping-pongサイクルの存在を実験的にしっかり示すことは困難でした。
勝間准教授のグループは、2014年にカイコにおいて雌化を担うFem piRNAとそのターゲットであるMasc遺伝子を同定しています(http://www.a.u-tokyo.ac.jp/topics/2014/20140515-1.html)。 このFem piRNAとMasc mRNAから産生されるMasc piRNAは、ping-pongサイクルによって増幅するペアの条件を満たしていました。また、反復配列から産生される大部分のping-pong piRNAとは異なり、Fem piRNAとMasc piRNAは1:1に対応するpiRNAペアでした。
そこで、このFem piRNAとMasc piRNAを利用して、人工的にping-pong piRNAを産生する実験系の構築に挑みました。
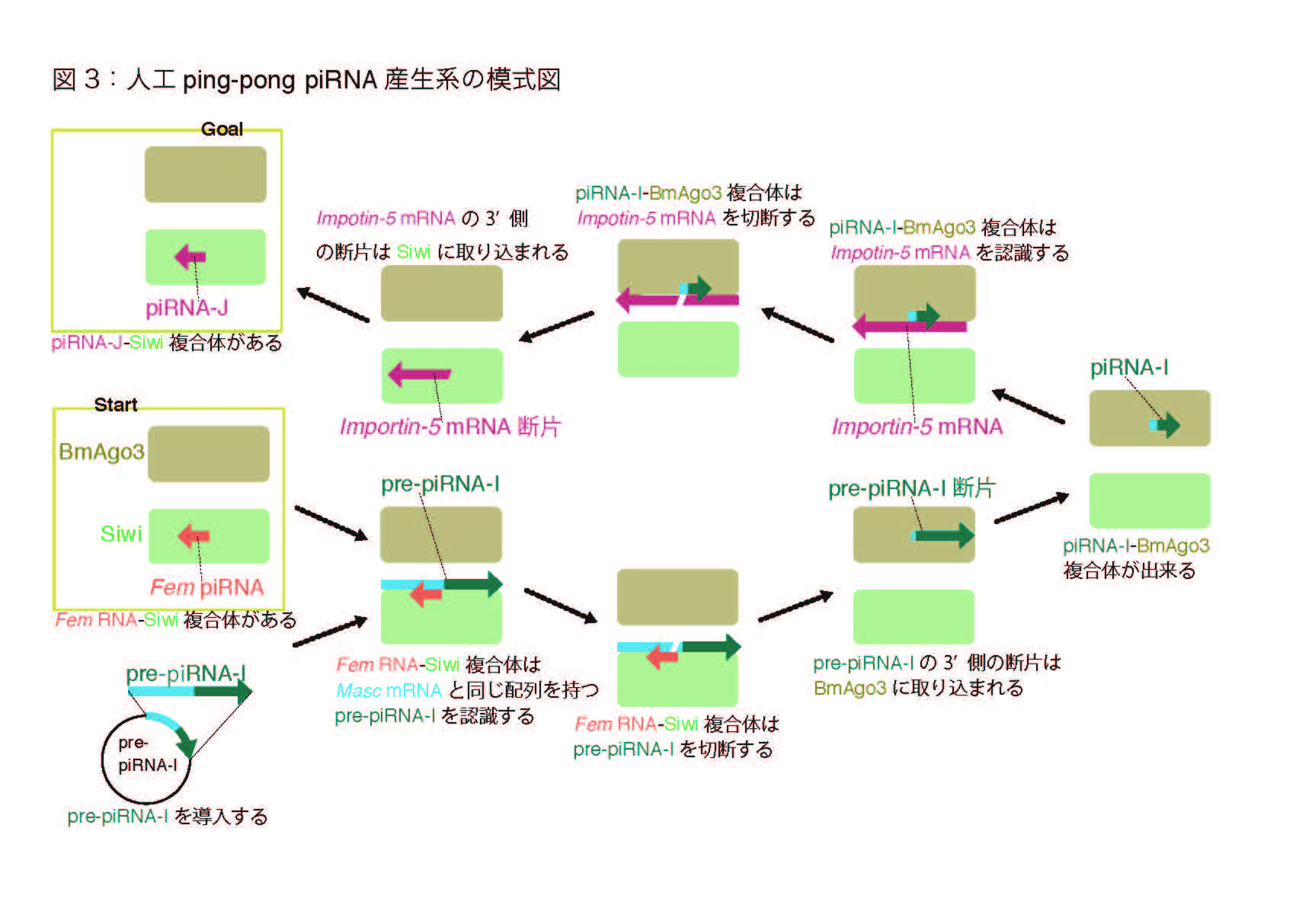
まず、人工的に設計したpiRNA前駆体(pre-piRNA-I:piRNA-Iは内在性の遺伝子であるImportin-5をターゲットにするよう設計されている)を発現するプラスミドを構築し、カイコ培養細胞BmN-4に導入しました。BmN-4細胞はping-pongサイクルに必要な2種類のPIWIタンパク質(Siwi及びBmAgo3)を発現し、それぞれに結合するpiRNAがお互いping-pong パートナーとしての特徴を持つことが知られている培養細胞です(図2)(Kawaoka et al., RNA, 2009 )。pre-piRNA-IはFem piRNA-Siwi複合体に認識される配列を持つため、成熟型のpiRNAであるpiRNA-Iを産生しました。さらに、piRNA-Iはもう一方のPIWIタンパク質であるBmAgo3と細胞内で複合体を作ることが確認できました。このpiRNA-I-BmAgo3複合体はBmN-4細胞で発現するImportin-5を認識/切断することが出来るため、piRNA-Jという人工的なpiRNAを産生すると予想出来ます(図3)。実際、次世代シークエンサーとインフォマティクスを用いた解析から、pre-piRNA-Iを導入した細胞ではpiRNA-Jが産生され、Siwiと結合していることがわかりました。
この結果から、Siwi-piRNA複合体によるpiRNA産生経路(ping)と、そのpiRNAとBmAgo3との複合体がpiRNAを産生し、もう一方のPIWIタンパク質
であるSiwiと結合する経路(pong)が連続的に働いていることが証明できました。
一方、本実験系を応用することで、piRNAがターゲット配列を認識するのに必要な配列の条件を探索しました。piRNA-IによるImportin-5の認識の程度は、piRNA-Jの産生量に対応すると考えられます。そこでpiRNA-Iと
Importin-5の配列の相補性が異なるようなpiRNA前駆体を設計し、それらをBmN-4細胞に導入した際のpiRNA-Jの産生量を調査しました。その結果、5’末端から17塩基分連続した相補性があれば、piRNAによってターゲット配列を認識できること、および22塩基分の相補性があれば、それ以上の相補性は認識効率には影響しないことを明らかにしました。
【本研究の将来】
本研究成果は、動物に広く保存された免疫システムであるpiRNA経路の中核を担うping-pongサイクルの存在を実験的に示した研究です。また、本研究で構築した人工ping-pong piRNA産生系を用いることで、piRNAの配列に隠された重要な特徴を明らかにすることできると期待されます。
本研究は、農林水産省のイノベーション創出基礎的研究推進事業「チョウ目昆虫における性操作技術の開発」(平成26年度より農林水産業・食品 産業科学技術研究推進事業に移管)、および文部科学省科学研究費補助金(基盤研究(A)、基盤研究(B)特設分野、特別研究員奨励費)による支援を受けて行われました。
発表雑誌
- 雑誌名
- 「RNA」(オンライン版の場合10月24日)
- 論文タイトル
- Artificial “ping-pong” cascade of PIWI-interacting RNA in silkworm cells
- 著者
- Keisuke Shoji, Yutaka Suzuki, Sumio Sugano, Toru Shimada, and Susumu Katsuma
- DOI番号
- 10.1261/rna.058875.116
- 論文URL
- http://rnajournal.cshlp.org/content/early/2016/10/24/rna.058875.116.abstract
問い合わせ先
東京大学大学院農学生命科学研究科 生産・環境生物学専攻 昆虫遺伝研究室
准教授 勝間 進(かつま すすむ)
Tel:03-5841-8994
Fax:03-5841-8993
Mail:katsuma@ss.ab.a.u-tokyo.ac.jp
用語解説
- 注1 PIWI-interactingRNA (piRNA)
- 小分子RNAの一つであり、生殖組織(細胞)に多く存在する。 RNA切断活性を持つPIWIタンパク質と結合して、相補的な配列を持つ利己的遺伝子(トランスポゾンなど)由来のRNAを切断する。
- 注2 ping-pong サイクル
- piRNA産生経路の一つ であり、2つの異なるPIWIタンパク質が関与する(カイコの場合はSiwiとBmAgo3)。カイコの場合、piRNAとSiwiの複合体がトランスポゾンmRNAを切断すると、切断された方は異なるPIWIタンパク質であるBmAgo3と結合する。その後、成熟型のpiRNAとなり、BmAgo3との複合体がもとのpiRNA前駆体となるトランスポゾンmRNAを切断する。このサイクルが繰り返されることで、トランスポゾンmRNAが分解され、それと同時にpiRNAの産生も行われる(図1参照)