植物が高温や乾燥などの劣悪なストレスに素早く応答するしくみを発見 ~高温・乾燥ストレス応答性転写因子DREB2Aのストレスによる活性化機構の解明~
- 発表者
-
溝井 順哉(東京大学大学院農学生命科学研究科応用生命化学専攻 准教授)
金澤 夏美(東京大学大学院農学生命科学研究科応用生命化学専攻 修士2年:研究当時)
城所 聡(東京大学大学院農学生命科学研究科応用生命化学専攻 助教)
高橋 史憲(理化学研究所 環境資源科学研究センター 機能開発研究グループ 研究員)
秦 峰(国際農林水産業研究センター 研究員:研究当時)
森本 恭子(東京大学大学院農学生命科学研究科応用生命化学専攻 博士3年:研究当時)
篠崎 一雄(理化学研究所 環境資源科学研究センター 機能開発研究グループ グループディレクター)
篠崎 和子(東京大学大学院農学生命科学研究科応用生命化学専攻 教授)
発表のポイント
- 植物の高温と乾燥の両方のストレスに応答するマスター転写因子であるDREB2Aがストレス時に活性化する分子メカニズムを明らかにしました。
- DREB2Aの活性化の分子メカニズムが明らかになったことで、植物の高温や乾燥ストレス時の遺伝子発現制御の初期過程が解明されました。
- 活性化に関するアミノ酸配列をゲノム編集技術等により改変することで、作物の高温や乾燥耐性を向上させることができると期待されます。また、本研究を足掛かりに、植物の高温や乾燥ストレス感知機構の解明につながると期待されます。
発表概要
東京大学、理化学研究所、国際農林水産業研究センターの共同研究グループは、シロイヌナズナ(注1)のストレス応答で働くマスター転写因子(注2)の一つDREB2Aがストレスに応答して活性化する仕組みを明らかにしました。DREB2Aは、植物が高温や乾燥による劣悪なストレス条件にさらされたときに、これらのストレスに耐えるための一群の遺伝子を発現させる機能をもちますが、通常生育条件では内部に存在するNRDと名付けられた配列の働きで分解され、不活性化されています。このことから、植物がストレスを受けるとNRDが働かなくなり、その結果DREB2Aが活性化すると考えられていましたが、その仕組みは明らかになっていませんでした。
本研究では、通常生育条件ではDREB2AのNRDがリン酸化(注3)され、このリン酸化により分解系に認識されることが明らかになりました。さらに高温ストレスに応答してこのリン酸化が抑制され、その結果DREB2Aが活性化するという仕組みが解明されました(図1)。NRDのリン酸化による活性制御は、常に変化するストレス環境への素早い応答に役立っていると考えられ、今後、このリン酸化を制御する仕組みを明らかにしていくことで、植物の高温および乾燥ストレスの感知機構の解明につながると考えられます。また、NRDに類似した配列は、作物を含む多くの植物で共通しているため、NRDにあたる配列をゲノム編集等によって改変することにより、植物の高温や乾燥耐性の向上につなげることができると考えられます。
発表内容
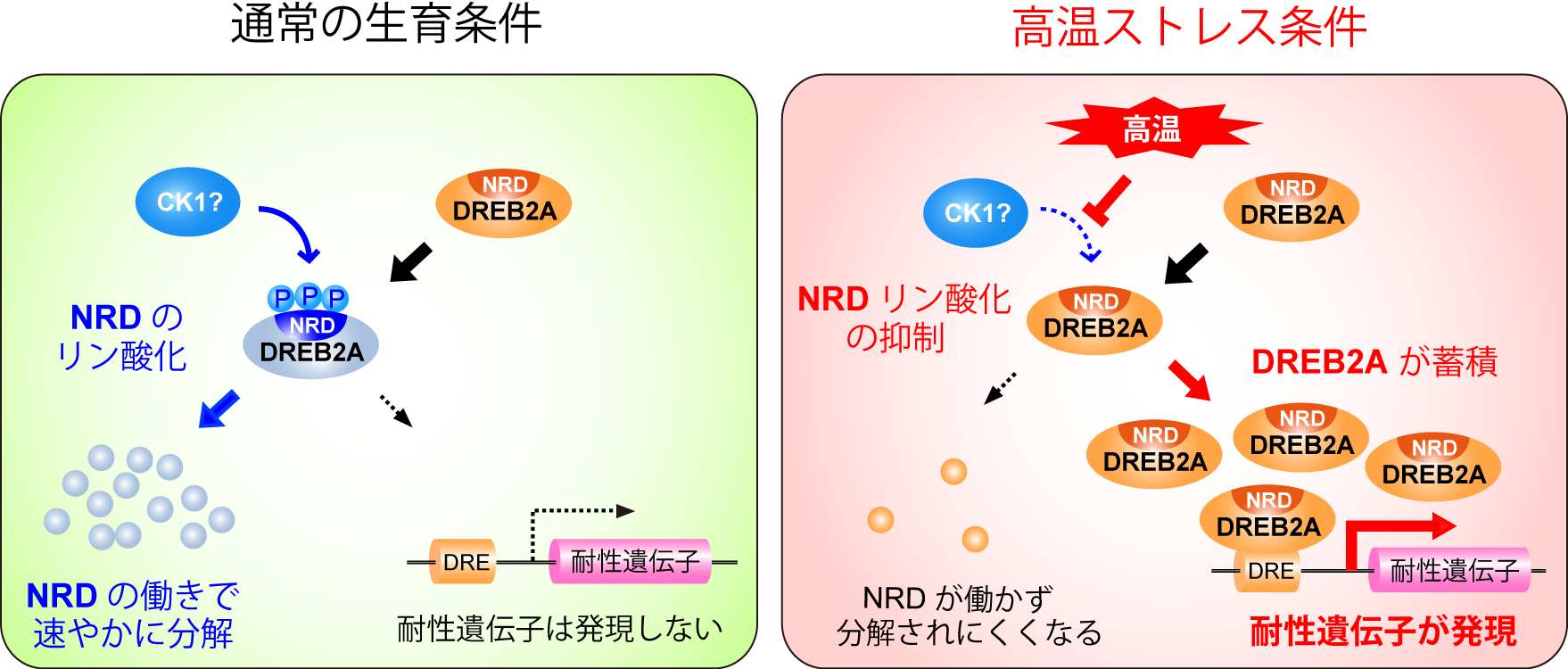
図1 シロイヌナズナの高温・乾燥ストレス応答で機能する転写因子DREB2Aは通常時は分解されていますが、ストレスを受けると分解が抑制されて蓄積し、耐性遺伝子の発現を活性化することができます。本研究で、通常の生育条件ではDREB2Aの内部にあるNRD配列がリン酸化され、このリン酸化が分解を促進することが明らかになりました(左)。さらに高温ストレスを受けた植物では、このリン酸化が阻害されることが明らかになりました(右)。その結果、分解されにくくなったDREB2Aは核内に蓄積し、耐性遺伝子の発現を活性化することができるようになります。
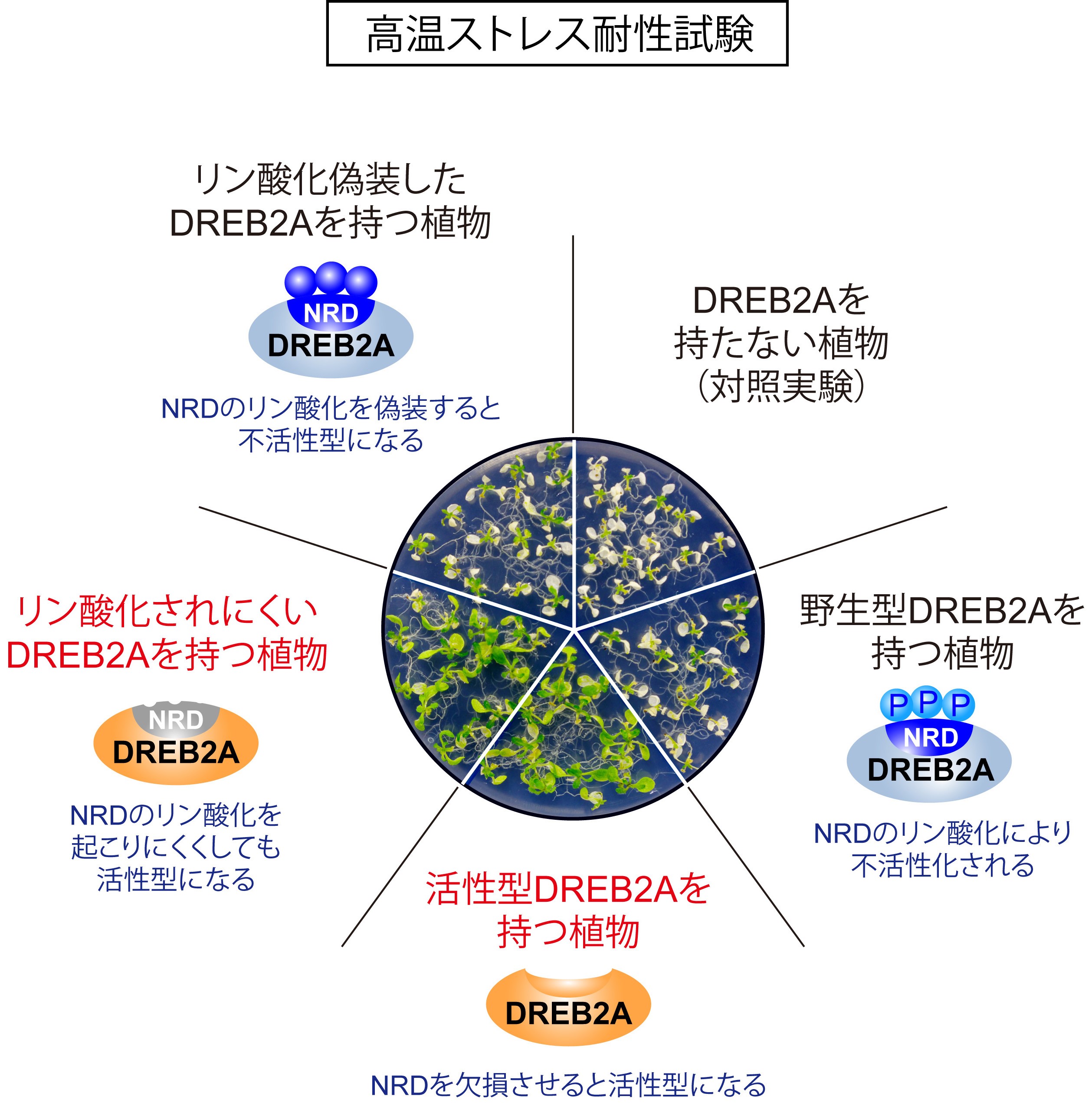
図2 NRDを欠損させた活性型DREB2Aは、野生型DREB2Aと比べて、より強い高温ストレス耐性を植物に付与することができます。NRDの配列を改変してリン酸化されにくいようにしたDREB2Aも、活性型DREB2Aと同様の強いストレス耐性を植物に付与することができました。一方で、NRDの配列を改変してリン酸化を偽装するようにしたDREB2Aは、高温ストレス耐性を付与することができませんでした。
近年、気候変動の影響で世界的に干ばつや熱波による作物の被害が頻繁に起きるようになってきています。干ばつや熱波により、植物は乾燥や高温といった生育や生存に大きく影響する環境ストレスを受けますが、その一方で、植物は自ら動くことができない代わりに、様々な環境ストレスに応答して、より強いストレスに耐えるように自らを変えていくことができます。とりわけ、ストレス耐性を向上させる遺伝子を発現させることは、耐性獲得の決定的なステップであることから、その仕組みを解明することは、干ばつや熱波の影響を受けにくい植物の開発にとって重要です。
シロイヌナズナのDREB2Aは高温や乾燥ストレスに素早く応答し、耐性遺伝子を発現させる働きを持つマスター転写因子です。一方でDREB2Aが働くと、ストレス耐性が向上すると同時に生育も阻害されるため、通常の生育条件では、DREB2Aは作られても速やかに分解され、機能しないようになっています。そこで、植物が高温や乾燥ストレスを受けるとDREB2Aの分解を抑制し活性化する仕組みが存在すると考えられていましたが、その仕組みはこれまで不明でした。
DREB2Aの分解は、内部にあるNRDとよばれる30アミノ酸程度からなる配列により促進されています。様々な植物のゲノム情報に基づいて配列を比較したところ、多くの植物のDREB2A相同タンパク質がNRDと類似したアミノ酸配列を持っていて、特にセリンおよびトレオニン残基が連続した配列が保存されていることが見いだされました。セリンおよびトレオニン残基は、リン酸化による修飾を受けることから、これらの残基をリン酸化されないアラニンあるいはリン酸化状態を偽装するアスパラギン酸に置換したDREB2Aをつくり、植物体内での働きを比較しました。その結果、アラニンに置換した場合には、NRDを取り除いた恒常的活性型DREB2A (DREB2A CA)と同じようにDREB2Aの分解が抑制されて活性が上昇し、植物の高温耐性も向上しました(図2)。一方で、アスパラギン酸に置換した場合には、DREB2Aの分解は抑制されず、植物の高温耐性も野生型と同様でした。実際に通常生育条件で生育させた植物内でDREB2Aのリン酸化状態を調べたところ、大部分のDREB2Aはリン酸化されていること、またリン酸化型DREB2Aは分解されやすいことが明らかになりました。対照的にNRDを持たず分解されにくいDREB2A CAは、その大部分がリン酸化されていませんでした。さらに、カゼインキナーゼ1(CK1)というタンパク質リン酸化酵素の働きを薬剤で阻害したところ、通常条件下でもDREB2Aのリン酸化が起こらなくなり、分解も抑制されました。以上の結果から、通常生育条件下ではDREB2Aが合成されると速やかにNRDがリン酸化を受け、このリン酸化によりDREB2Aの分解が促進されることが明らかになりました(図1左)。
植物が高温ストレスを受けると、DREB2Aは分解が抑制されて活性化します。そこで、この時のDREB2Aのリン酸化状態を調べました。その結果、高温ストレス処理によって大量に蓄積するDREB2Aの大部分はリン酸化されていないこと、またその原因はDREB2Aのリン酸化が阻害されることであることが明らかになりました。以上の実験から、植物が高温ストレスを受けると、そのストレスのシグナルがNRDのリン酸化の抑制という形でDREB2Aに伝わり、その結果DREB2Aの分解が抑制されて大量に蓄積するという活性化の仕組みが明らかになりました(図1右)。
高温や乾燥によるストレス状態は、日照、気温、湿度、土壌水分、風速など、様々な環境条件によって刻々と変化するため、植物はその時点のストレス状態に素早く対応していく必要があります。今回明らかにしたようなリン酸化による活性の調節は短時間でおこるため、環境条件の変化に対応し、耐性と成長のバランスを保つために有利に働くと考えられます。今後、DREB2Aのリン酸化がストレスによって抑制される仕組みを明らかにすることで、植物がストレスをどのように感じ取っているのかを明らかにしていくことができます。また、NRDに類似した配列は、多くの作物のDREB2A相同タンパク質にも共通しています。今回、NRDへのアミノ酸置換により、シロイヌナズナのストレス耐性を変化させることができました(図2)。したがって、この配列およびリン酸化の仕組みを標的とした遺伝子改変により作物のストレス耐性を変化させることが期待できます。
発表雑誌
- 雑誌名
- 「Journal of Biological Chemistry」November 28, 2018
- 論文タイトル
- Heat-induced inhibition of phosphorylation of the stress-protective transcription factor DREB2A promotes thermotolerance of Arabidopsis thaliana
- 著者
- Junya Mizoi, Natsumi Kanazawa, Satoshi Kidokoro, Fuminori Takahashi, Feng Qin, Kyoko Morimoto, Kazuo Shinozaki, Kazuko Yamaguchi-Shinozaki*
- DOI番号
- 10.1074/jbc.RA118.002662
- 論文URL
- http://www.jbc.org/content/early/2018/11/28/jbc.RA118.002662.abstract
問い合わせ先
東京大学大学院農学生命科学研究科応用生命化学専攻 植物分子生理学研究室
准教授 溝井 順哉(みぞい じゅんや)
Tel:03-5841-8111
Fax:03-5841-8009
E-mail:ajmizoi<アット>mail.ecc.u-tokyo.ac.jp <アット>を@に変えてください。
東京大学大学院農学生命科学研究科応用生命化学専攻 植物分子生理学研究室
教授 篠崎 和子(しのざき かずこ)
Tel:03-5841-8137
Fax:03-5841-8009
E-mail:akys<アット>mail.ecc.u-tokyo.ac.jp <アット>を@に変えてください。
用語解説
- 注1 シロイヌナズナ
- 植物科学の研究で使われるモデル生物で、学名はArabidopsis thaliana。植物体の小ささや世代時間の短さなど様々な利点から、世界中で使われていて、突然変異体やデータベースなどの研究資源も充実しています。環境ストレスに耐える能力が強いため、ストレス応答の研究にも適した実験材料でもあります。
- 注2 転写因子
- 遺伝子が機能するためには、DNA配列にコードされた遺伝子情報を鋳型にしてmRNAが合成される必要があります(転写)。転写因子は、遺伝子近傍にある特有の配列を認識して結合し、mRNA合成のタイミングと合成量を決定する働きを持っています。一般的に一つの転写因子は複数の遺伝子を同時に制御できますが、中でも多くの遺伝子の発現を制御し、細胞の応答や発生の方向性を決定づけるような転写因子は、マスター転写因子と呼ばれることがあります。
- 注3 リン酸化
- タンパク質の機能の調節にかかわる主要な翻訳後修飾機構の一つ。リン酸化は多くの場合可逆的で、新たな遺伝子発現を必要とせずタンパク質の性質を変えることができるので、既に合成されたタンパク質の機能を素早く変化させるスイッチとして働くことができます。