植物レクチンがウイルス増殖阻害剤として利用できる可能性を発見
- 発表者
- 吉田 哲也(東京大学大学院農学生命科学研究科 生産・環境生物学専攻 博士課程3年生)
白石 拓也(東京大学大学院農学生命科学研究科 生産・環境生物学専攻 博士課程;研究当時)
岡野 夕香里(東京大学大学院農学生命科学研究科 生産・環境生物学専攻 特任研究員)
藤本 祐司(東京大学大学院農学生命科学研究科 生産・環境生物学専攻 修士課程2年生)
遊佐 礼(東京大学大学院農学生命科学研究科 生産・環境生物学専攻 博士課程3年生)
前島 健作(東京大学大学院農学生命科学研究科 生産・環境生物学専攻 助教)
山次 康幸(東京大学大学院農学生命科学研究科 生産・環境生物学専攻 准教授)
発表のポイント
- レクチン型抵抗性遺伝子JAX1による植物ウイルス感染阻害メカニズムを明らかにしました。
- JAX1が「ウイルス複製工場」に侵入し、ウイルスの増殖を阻害することを解明しました。
- 耐性ウイルスが発生しにくい植物ウイルス阻害剤開発につながることが期待されます。
発表概要
植物ウイルスはさまざまな農作物に感染し品質や収量に甚大な被害をもたらしますが、有効な化学農薬がなく防除が困難です。植物が持つウイルス抵抗性遺伝子の利用は最も有効な防除手段であり環境負荷も小さいため、抵抗性遺伝子の探索とそのメカニズムの解明が強く求められています。
2012年、東京大学大学院農学生命科学研究科の山次康幸准教授らの研究チームは、世界中の農作物に甚大な被害を与えている約40種類の「ポテックスウイルスグループ」(注1)の増殖を強力に抑える超広域抵抗性遺伝子JAX1(注2)を発見しましたが、この遺伝子は既存の植物ウイルス抵抗性遺伝子とは全く構造が異なる「レクチン」(注3)をコードしており、その抵抗性メカニズムは不明でした。
今回、同研究チームはポテックスウイルスの無細胞翻訳・複製系(注4)を構築して調べたところ、ウイルスが複製酵素を翻訳するとただちにJAX1が複製酵素と結合し、「ウイルス複製工場」(注5)に一緒に侵入して、ウイルスゲノムの複製を阻害することを明らかにしました。この新たな抵抗性メカニズムはウイルスの複製酵素が標的としており、JAX1を利用すれば防除戦略上で重要な多数の植物ウイルスに強力な抵抗性を付与できることが期待されます。
発表内容
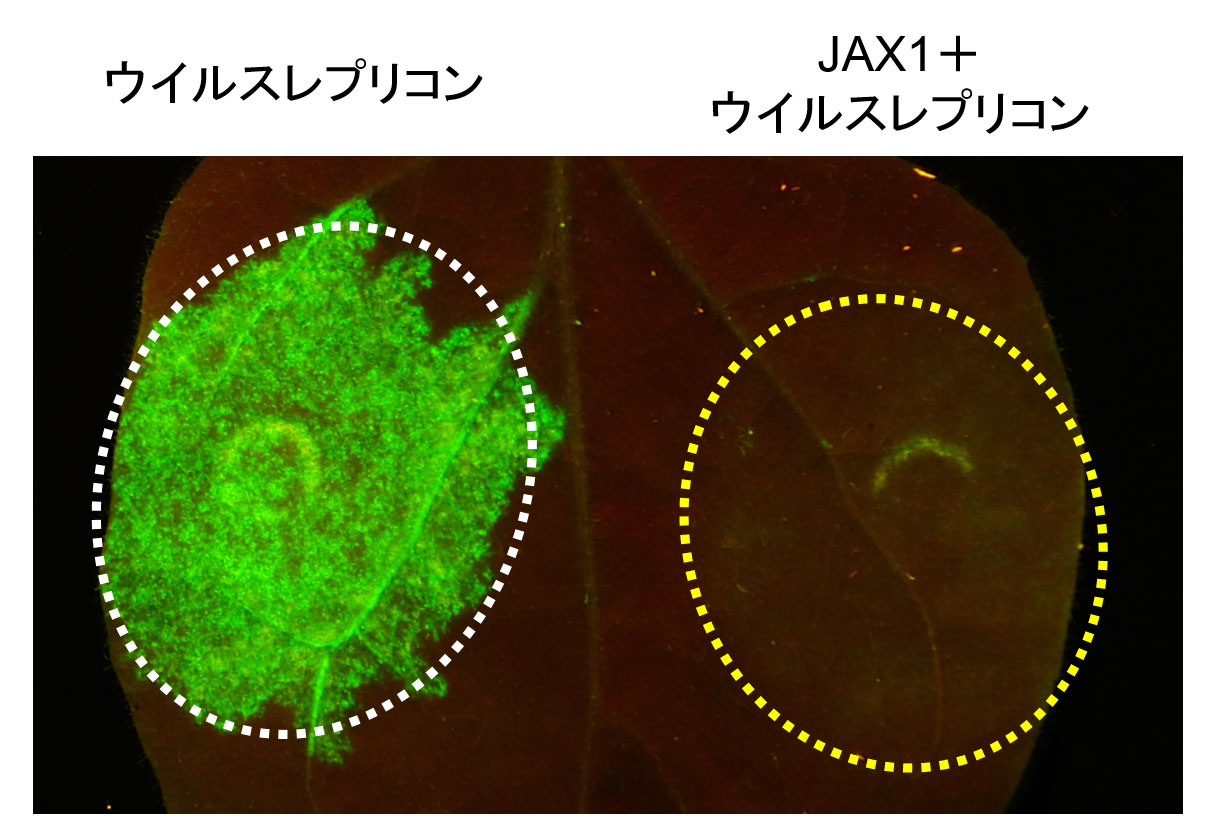
図1 JAX1によるウイルス複製阻害
隣接細胞への移行能を欠き、複製のみを行うウイルスレプリコンを植物に接種するとゲノム複製を行い,ウイルスに組み込んだGFPの蛍光を発する(左)が、JAX1を同時に発現させるとウイルスの複製が強く抑制され、GFPが観察されない(右)。
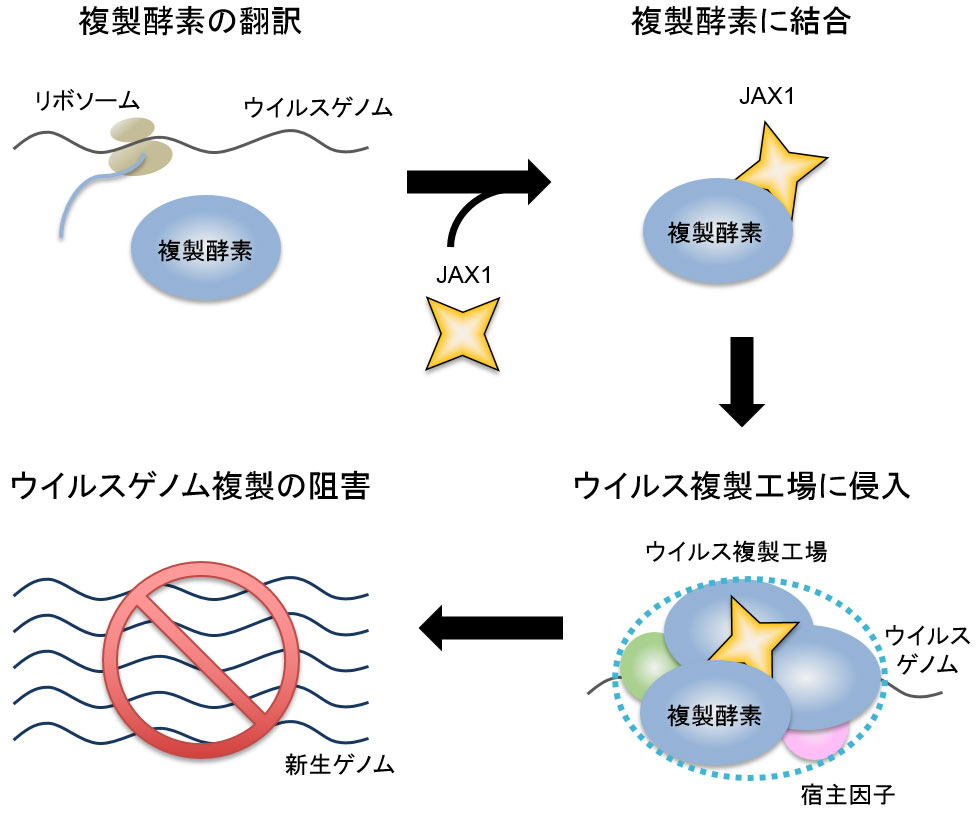
図2 JAX1によるウイルス複製阻害のモデル
JAX1は翻訳直後のウイルス複製酵素に結合し、ウイルス複製工場に侵入することによってウイルスゲノム複製を阻害する。
【研究の背景】
植物ウイルスは、多くの農作物に感染し多大な被害をもたらすため、その防除法の開発が、世界中でしのぎを削って試みられています。一般に、ウイルス抵抗性を示す植物品種(抵抗性品種)が育種され利用されますが、利用可能な抵抗性品種の種類は極めて限られています。そのため、抵抗性品種を用いた植物ウイルス防除においては、新たなウイルス抵抗性遺伝子の発見と、その機能の解明が非常に重要となります。ウイルス抵抗性品種の大部分は真正抵抗性(注6)とよばれる抵抗性によるもので、ウイルスに対する受容体として機能する「nucleotide-binding leucine-rich repeat(NLR)型抵抗性遺伝子(注6)」を持つことが知られており、その抵抗性誘導メカニズムが盛んに研究されてきました。一方、近年NLR型抵抗性遺伝子とは構造の異なるウイルス抵抗性遺伝子(non-NLR型抵抗性遺伝子(注7))が存在し、真正抵抗性とは異なる特徴を示すことが明らかになり、その機能に大きな注目が集まっています。しかし、non-NLR型抵抗性遺伝子による抵抗性メカニズムの多くは明らかにされておらず、解明が待たれています。
東京大学大学院農学生命科学研究科の山次康幸准教授らの研究チームは、世界中でさまざまな農作物に甚大な被害をもたらしている植物ウイルスグループである「ポテックスウイルスグループ」の感染を強力に阻害するnon-NLR型抵抗性遺伝子JAX1を2012年に発見しました。JAX1は糖鎖に結合する性質を持つタンパク質群「レクチン」の一種をコードするユニークな抵抗性遺伝子です。JAX1はポテックスウイルスの増殖を単細胞レベルで阻害することが示されていましたが、抵抗性メカニズムの詳細は不明でした。
【研究内容】
今回、同研究チームは、JAX1による抵抗性が発揮されるタイミングとその標的分子を解析しました。まず、植物体内で細胞内でのゲノム複製を行うウイルスレプリコン(注8)と同時にJAX1を発現させたところ、レプリコンの増殖が強く抑制されたことから、ウイルスのゲノム複製が阻害されると考えられました。続いて、JAX1がどのタイミングでウイルスゲノム複製を阻害しているのかを無細胞ウイルス翻訳・複製系を用いて解析したところ、JAX1がウイルス複製酵素の翻訳直後の極めて早いタイミングで作用し、ウイルスゲノムの複製を阻害することが明らかとなりました。この際のJAX1の存在部位を調べたところ、1,000 kDa以上のサイズの巨大なタンパク質複合体である「ウイルス複製工場」からJAX1が検出されました。また、JAX1がウイルス複製酵素と結合することも明らかにしました。これらの結果から、JAX1はウイルスの極めて初期の感染過程で複製酵素と結合することによりウイルス複製工場内部に侵入し、ウイルスのゲノム複製を阻害することが明らかとなりました。
【社会的意義・今後の展望など】
植物病害に対する抵抗性品種開発においては、耐性菌や耐性ウイルス(注9)の出現が最大の問題です。これまでの研究で、JAX1を利用した植物においては、耐性ウイルスが発生しにくいことが明らかになっていましたが、そのしくみは不明でした。今回JAX1がポテックスウイルスの複製を非常に早い段階で阻害することが明らかになりました。このことから、JAX1が複製の際にウイルスゲノムが突然変異して、耐性ウイルスが出現することを防いでいるものと考えられます。これは、ウイルス防除法として極めて有用です。
NLR型抵抗性遺伝子は遠縁な植物に導入すると正常に抵抗性を発揮しないことが多く、実用上の問題となっています。これは、NLR型抵抗性が細胞内で複雑なシグナル伝達経路を必要とする一方で、そのような経路が異なる植物の間では保存されていないことが一因と考えられています。今回、JAX1が単独でウイルスの複製工場に侵入し、ゲノム複製を阻害することが明らかとなりました。このことは、JAX1抵抗性がNLR型抵抗性とは異なり、細胞内の複雑なシグナル伝達経路を必要としないことを示しており、幅広い植物でJAX1を利用したウイルス防除が可能であることを示しています。
今回示されたJAX1による抵抗性メカニズムは、植物が持つnon-NLR型抵抗性遺伝子によるウイルス抵抗性メカニズムの多様性の一端を示す成果と考えられます。植物には、私達の知らないウイルス抵抗性メカニズムがまだ多く眠っているはずです。未知のnon-NLR型抵抗性遺伝子の探索と機能解析がさらに進めば、新たなウイルス抵抗性メカニズムの発見と、新たな防除戦略の開発が期待されます。
本研究は日本学術振興会科学研究費補助金の支援を受けて行われました。
発表雑誌
- 雑誌名
- Journal of Virology(1月17日オンライン公開予定)
- 論文タイトル
- The Plant Noncanonical Antiviral Resistance Protein JAX1 Inhibits Potexviral Replication by Targeting the Viral RNA-Dependent RNA Polymerase
- 著者
- Tetsuya Yoshida, Takuya Shiraishi, Yuka Hagiwara-Komoda, Ken Komatsu, Kensaku Maejima, Yukari Okano, Yuji Fujimoto, Akira Yusa, Yasuyuki Yamaji, Shigetou Namba*
- DOI番号
- 10.1128/JVI.01506-18
- 論文URL
- https://doi.org/10.1128/JVI.01506-18
問い合わせ先
東京大学大学院農学生命科学研究科 生産・環境生物学専攻 植物病理学研究室
准教授 山次 康幸(やまじ やすゆき)
Tel:03-5841-5092
Fax: 03-5841-5090
E-mail:ayyamaji<アット>mail.ecc.u-tokyo.ac.jp <アット>を@に変えてください。
研究室URL:http://park.itc.u-tokyo.ac.jp/ae-b/planpath/
用語解説
- 注1 ポテックスウイルスグループ
プラス1本鎖RNAをゲノムとする植物ウイルスグループで、分類学的にはポテックスウイルス属とよばれます。38種のウイルスが含まれ、ウイルス粒子は幅13 nm(ナノメートル)、長さ470–580 nmからなるひも状です。代表的なウイルスとしては、それぞれジャガイモ、トマト、ラン、ユリなどに感染し世界中で被害をもたらしているpotato virus X、pepino mosaic virus、cymbidium mosaic virus、plantago asiatica mosaic virusなどが含まれます。 - 注2 JAX1
モデル植物シロイヌナズナから発見されたポテックスウイルス抵抗性遺伝子で、ジャカリンというレクチンの一種に似たタンパク質をコードしています。これまでの解析により、JAX1は一般的な抵抗性遺伝子とは異なり、広範なウイルス(ポテックスウイルス全般)に抵抗性を発揮すること、細胞レベルでウイルスの増殖を阻害することなどが示唆されていましたが、その抵抗性メカニズムは不明でした。 - 注3 レクチン
糖鎖に結合するタンパク質の総称で、自然界に広く存在しています。植物には多くのレクチンが含まれており、初めて同定されたレクチンも植物に由来するものです。動物においては、病原体表面の糖鎖を認識することによって免疫系を活性化することなどが知られています。しかし、植物におけるレクチンの機能はよくわかっていません。 - 注4 無細胞ウイルス翻訳・複製系
試験管内でウイルスの翻訳(タンパク質の合成)および複製(新生ゲノムの合成)を再構成する実験系のこと。細胞壁や液胞の除去などのさまざまな処理を施した植物細胞を磨砕して得られた液中にはウイルスの翻訳や複製に必要な細胞内のさまざまな因子が保持されているため、この液にウイルスのゲノムを加えることで翻訳・複製過程を再構成し、そのメカニズムを詳細に解析することができます。本研究では、タバコに由来する培養細胞の抽出液(Komoda et al., PNAS 101:1863–1867)を用いることで、ポテックスウイルスで初めて本実験系の構築に成功しました。 - 注5 ウイルス複製工場
植物プラス鎖RNAウイルスは、植物細胞内で複製酵素を翻訳した後に、複製酵素、ウイルスRNAや宿主タンパク質を含む巨大なタンパク質・RNA複合体を形成し、この複合体が植物細胞の生体膜に結合することにより、複合体内部で新たなウイルスゲノムが合成されることから、ウイルス複製工場とよばれます。 - 注6 真正抵抗性とNLR型抵抗性遺伝子
真正抵抗性は植物が病原体に対して示す抵抗性反応の1種であり、現在利用されているさまざまな作物の抵抗性品種の大部分はこの抵抗性反応により病原体の感染を防いでいます。垂直抵抗性ともよばれます。病原体由来のタンパク質などを植物の抵抗性遺伝子(NLR遺伝子)が認識することにより誘導され、活性酸素の発生・防御遺伝子の発現・抗菌物質(ファイトアレキシンなど)の生成による病原体の不活化や、過敏感細胞死などによる病原体の封じ込めを行います。 - 注7 non-NLR型抵抗性遺伝子
NLR型ウイルス抵抗性遺伝子とは構造の異なる抵抗性遺伝子。2000年に初めて報告されて以来、その報告数が近年徐々に増加しています。多様な種類のタンパク質をコードしており、それぞれが独自のメカニズムでウイルスに抵抗性を発揮すると想定されていますが、その多くは不明です。 - 注8 レプリコン
ウイルスゲノムのうち、隣接細胞への移行に必要な移行タンパク質に変異を導入することにより複製のみを行うようになったウイルス変異体で、ウイルスの複製の有無を判定する実験に用いられます。 - 注9 耐性菌・耐性ウイルス
ヒトの病気の治療には抗生物質が有効ですが、抗生物質が効かない耐性菌の発生が問題になっています。植物の病気も同様で、抵抗性品種に耐性菌や耐性ウイルスが発生し大きな被害を引き起こします。耐性ウイルスでは、抵抗性タンパク質が標的とするウイルスタンパク質に変異が起こり、抵抗性タンパク質の認識を回避することで抵抗性を免れています。