葉化病発症の仕組みを構造化学的に解明 ―葉化病治療薬開発や新品種開発に新たな道―
- 発表者
- 岩渕 望(東京大学大学院農学生命科学研究科 生産・環境生物学専攻 博士課程3年)
前島 健作(東京大学大学院農学生命科学研究科 生産・環境生物学専攻 助教)
北沢 優悟(東京大学大学院農学生命科学研究科 生産・環境生物学専攻 特任研究員)
山次 康幸(東京大学大学院農学生命科学研究科 生産・環境生物学専攻 教授)
発表のポイント
- 植物病原細菌ファイトプラズマが持つ花の葉化因子「ファイロジェン」の立体構造を解明しました。ファイトプラズマの病原性因子の立体構造解明は世界初です。
- ファイロジェンの立体構造は、植物の花形成因子同士の結合を担う保存領域と類似しており、この結合の仕組みを模倣し阻害することが、葉化病発症の普遍的な仕組みであることを明らかにしました。
- 本研究成果は、治療薬の開発や、付加価値の高い緑色の花や寿命の長い花卉品種などの作出に新たな道を拓くことが期待されます。
発表概要
ファイトプラズマ(注1)は様々な植物の花を葉に変える葉化病を引き起こす植物病原細菌です。近年、ファイトプラズマの分泌タンパク質「ファイロジェン」(注2)が葉化因子として特定され、その標的が植物の花形成因子(MADS転写因子)(注3)であることが明らかになっていましたが、ファイロジェンがどのように花形成因子を特異的に認識しているのかは不明でした。今回、東京大学大学院農学生命科学研究科の岩渕望大学院生と前島健作助教らの研究グループは、ファイロジェンの立体構造をX線結晶構造解析法により解明しました(図1)。ファイロジェンは植物の花形成因子と結合して分解する性質を持ちますが、ファイロジェンの構造が花形成因子同士の結合を担うKドメインの構造と類似しており、この構造に変異を加えるとファイロジェンの活性が失われることを特定しました(図2)。このことは、ファイロジェンが標的である植物の花形成因子の構造を模倣することで、幅広い植物の花形成因子同士の結合を阻害しつつ結合・分解してその働きを妨げ、花を葉に変化させていることを示しています。本研究成果は、ファイトプラズマの病原性因子の立体構造が解明された初めての事例です。
本研究成果は、葉化病発症の普遍的な仕組みを明らかにし治療薬の開発を可能にするとともに、ファイロジェンという1種類のタンパク質を利用するだけで、付加価値の高い葉化品種を様々な植物において作出できることを示す基盤的成果です。
発表内容
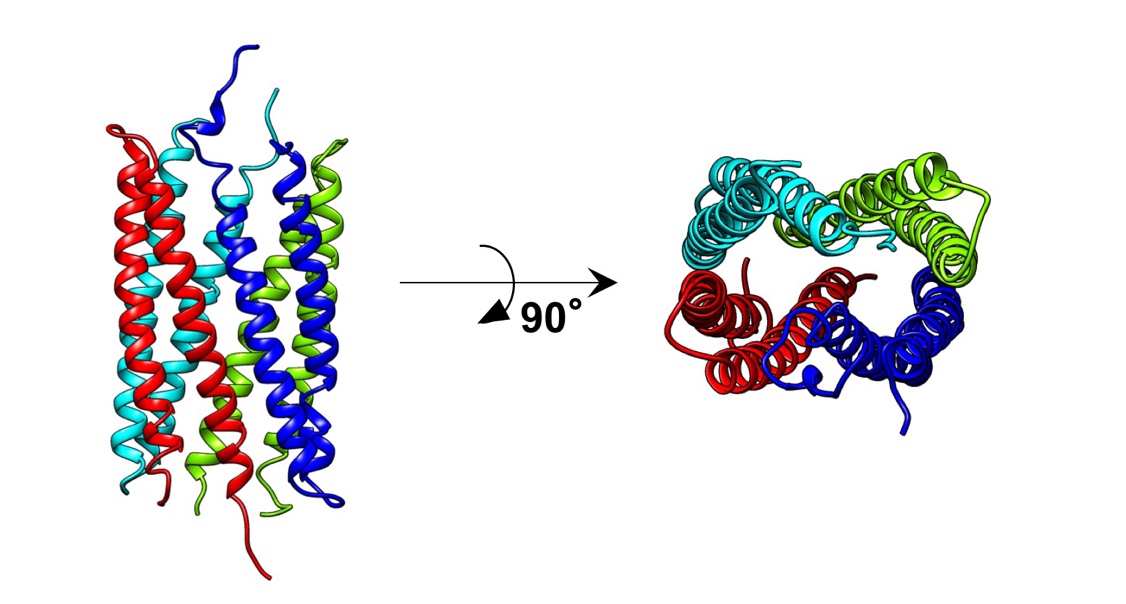
図1 葉化因子ファイロジェンの立体構造
X線結晶構造解析により得られた、ファイロジェン4分子から構成される結晶構造。
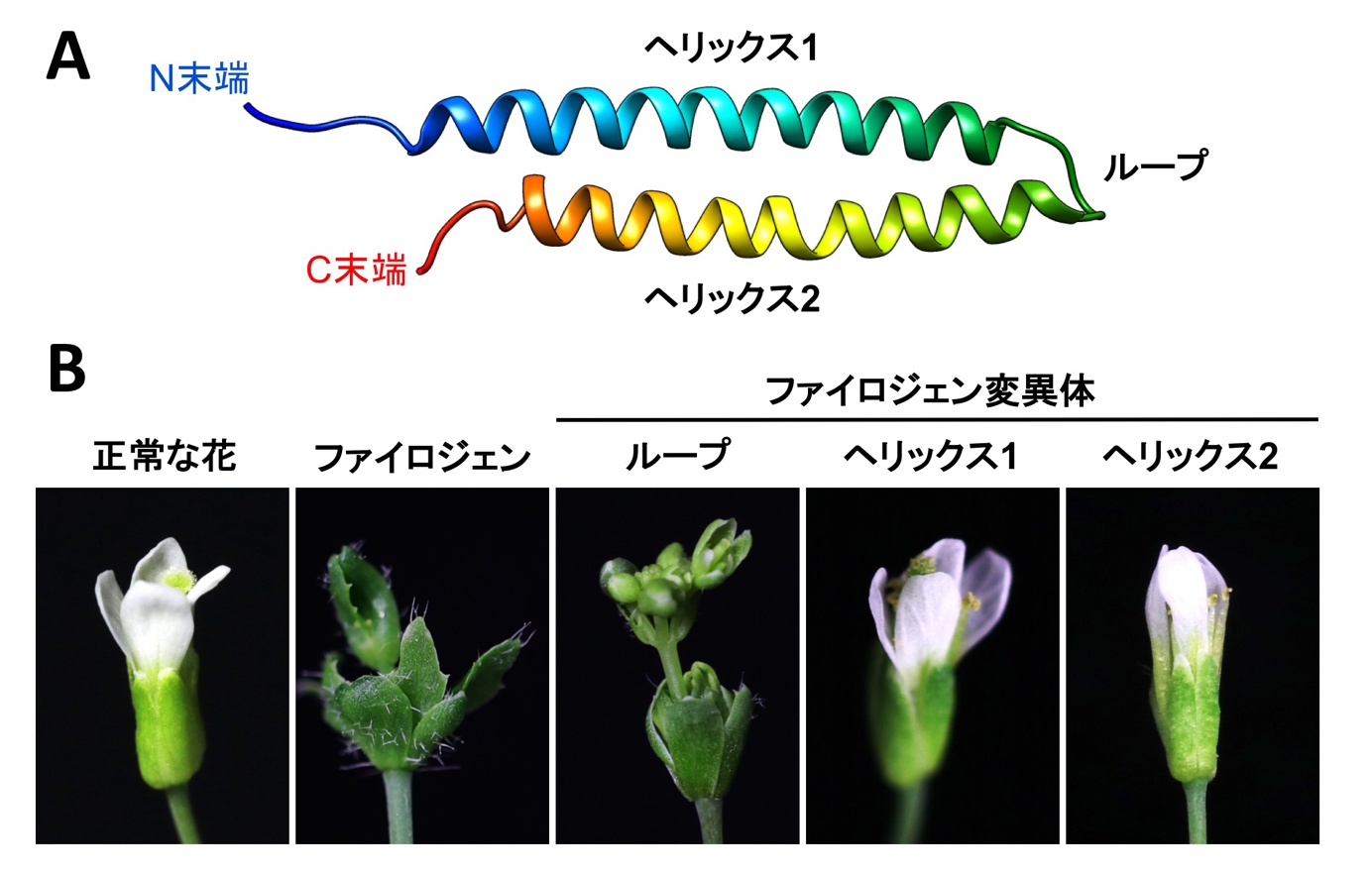
図2 ファイロジェンの構造と機能の関係
(A)ファイロジェンは2つのヘリックスが短いループで繋がれた構造をとる。(B)ループ部分を変異させても葉化能に影響しないが、どちらのヘリックスに変異を入れても、ファイロジェンの葉化能が失われる。
1.研究の背景
ファイトプラズマは、1967年に東京大学農学部の植物病理学研究室において発見された、植物の篩部に寄生しヨコバイなどの昆虫により媒介される植物病原細菌であり、葉化病や天狗巣病などを引き起こします。同研究室では、2004年の世界初のファイトプラズマのゲノム解読以降、ゲノム情報に基づき、葉化病や天狗巣病を始めとしたファイトプラズマによるさまざまな病徴の発生メカニズムを明らかにしてきました。
植物の花は葉が変化して形成されるものであり、がく、花びら、雄しべ、雌しべの各器官から構成されます。花の形成過程では、A・B・C・Eクラスに分類される花形成因子(MADS転写因子)が特定の組み合わせで多量体を形成することで転写因子として機能し、がくや花びらへと細胞が分化することが知られています。これまでの研究から、ファイトプラズマの葉化因子「ファイロジェン」がA・Eクラスの花形成因子に結合し、植物のプロテアソームを介して分解することにより花の形成過程を阻害し、葉化を誘導することが明らかになっていましたが、ファイロジェンがどのようにして特定の花形成因子を認識しているのかはわかっていませんでした。
2.研究内容
今回、同研究室の岩渕望大学院生と前島健作助教らは、X線結晶構造解析により、2.4Åの解像度でファイロジェンの立体構造の解明に成功しました。立体構造に基づくモデリング解析から、各種ファイトプラズマが持つファイロジェンは、2つの両親媒性ヘリックス(注4)からなる共通の立体構造を有することが明らかになりました。またこの構造は、花形成因子のKドメインの立体構造と類似していることを明らかにしました。Kドメインは花形成因子同士の特異的な結合を司ることから、ファイロジェンはKドメインを模倣した構造をとることにより、特定の花形成因子同士の結合を阻害しつつ分解へと導いていると考えられます。実際に、両親媒性ヘリックスに変異を入れると、花形成因子に対する結合・分解活性が失われるとともに、植物への葉化誘導能が失われました。
3.社会的意義
花が葉化すると、ファイトプラズマが寄生できる篩部組織が増大するとともに、植物の寿命が延びて若い枝葉が伸長し続けることから、媒介昆虫による吸汁を受けやすくなり、ファイトプラズマが新たな植物へと伝染するチャンスが高まります。このことは、幅広い植物を葉化させることが、ファイトプラズマにとって非常に重要な生存戦略であることを示しています。
花器官の形成には、きわめて複雑な遺伝子の発現制御が関わることが知られており、その制御を司っているのが花形成因子のKドメインを介した多量体化です。この多量体化の分子機構は植物間で高度に保存されていますが、本研究により、ファイロジェンはその分子機構を模倣し阻害することで、どんな植物においても花の葉化という普遍的な機能を発揮できることがわかってきました。ファイトプラズマはファイロジェンの獲得を通じて、どんな植物に感染した場合でも自身の生存の機会を最大限に向上させることができるようになったと考えられます。
ファイロジェンによる植物普遍的な葉化の仕組みの解明は、葉化病の治療薬のみならず、様々な植物における付加価値の高い葉化品種の作出や寿命の長い花卉品種の作出にも活用できると考えられ、今後の品種開発への貢献が期待されます。
発表雑誌
- 雑誌名
- Biochemical and Biophysical Research Communications(4月19日オンライン公開)
- 論文タイトル
- Crystal structure of phyllogen, a phyllody-inducing effector protein of phytoplasma
- 著者
- Nozomu Iwabuchi, Kensaku Maejima*, Yugo Kitazawa, Hideyuki Miyatake, Masanobu Nishikawa, Ryosuke Tokuda, Hiroaki Koinuma, Akio Miyazaki, Takamichi Nijo, Kenro Oshima, Yasuyuki Yamaji, Shigetou Namba
- DOI番号
- 10.1016/j.bbrc.2019.04.060
- 論文URL
- https://www.sciencedirect.com/science/article/pii/S0006291X19306886
問い合わせ先
東京大学大学院農学生命科学研究科 生産・環境生物学専攻 植物病理学研究室
助教 前島 健作 (まえじま けんさく)
Tel:03-5841-1613
Fax:03-5841-5090
E-mail:amaejima<アット>mail.ecc.u-tokyo.ac.jp <アット>を@に変えてください。
研究室URL:http://park.itc.u-tokyo.ac.jp/ae-b/planpath/
用語解説
- 注1 ファイトプラズマ
1967年にマイコプラズマ様微生物(mycoplasma-like organism, MLO)として発見され、2004年に全ゲノムが解読された、ファイトプラズマ属(モリキューテス綱)に分類される植物病原細菌。1,000種以上の植物に感染し、世界中の農業生産に被害をもたらしている重要な植物病原細菌である。細胞壁を欠いた直径0.1〜0.8 μmの不ぞろいな粒子状で、細菌の中でも最小である。植物の篩部に寄生し、ヨコバイ等の昆虫により植物から植物へと媒介される。植物に萎縮病、天狗巣病、葉化病(花の葉化・緑化・突き抜け)などの特徴的な病気を引き起こし、植物を枯らしてしまうことも多い。ファイトプラズマ病は、私たちの身近にも頻繁に認められる病気であり、葉化病によって緑色の花が咲くアジサイは商品価値が認められ、品種登録されていた例もある。また、クリスマスシーズンの風物詩である鉢植えのポインセチア(枝分れが豊富で、矮性化するタイプ)は、全て背丈を小さくするためにファイトプラズマに人工的に感染させられ、天狗巣病を発病したものであり、健全なポインセチアは2mにもなる。
<ファイトプラズマによって引き起こされる主な症状>
萎縮:茎や葉の生長が害され、著しく矮性となる症状
天狗巣:側芽が異常に発達し、小枝が密生する症状
花の葉化:花びらやがく・雌しべ・雄しべが葉に置き換わってしまうこと
- 注2 葉化因子「ファイロジェン」
「葉化病(phyllody、phyllo-(葉)+-ody(になる変化))」の原因である遺伝子ファミリー「phyllody-inducing gene family(葉化誘導遺伝子ファミリー)」の略。phyllo-(葉)+-gen(を生ずるもの=gene(遺伝子))。ファイロジェンは約90アミノ酸程度の小さなタンパク質で、ファイトプラズマ属全般に高度に保存されている。これまで、ファイロジェンがあらゆる植物のMADS転写因子を標的とすることが明らかとなっていたが、今回その仕組みが明らかになった。 - 注3 MADS転写因子
花の形成を制御する転写因子。どの植物細胞が花びらやがくになるかは、MADS転写因子の組み合わせによって決まる。4つのMADS転写因子が複合体(4量体)を形成して機能すると考えられており、「カルテットモデル」と呼ばれる。複合体形成にはMADS転写因子を構成する4つのドメインのうち、Kドメインが関わる。
カルテットモデルに関わるMADS転写因子はA・B・C・Eクラスに分類され、A・Eクラスの転写因子が発現した細胞ではA・A・E・Eの4量体が形成されがくになる。A・B・B・Eの組合せで花弁(花びら)、B・B・C・Eで雄しべ、C・C・E・Eで雌しべになる。これらの遺伝子が働かなくなった「変異体」では、がくが雌しべになったり、花びらががくになったりする。一般に見られる「八重咲き」品種の花はこれらの遺伝子が働かなくなったために雄しべや雌しべが花びらに変化したもので、こうした遺伝子の変異が育種的に固定されたものである。 - 注4 両親媒性ヘリックス
αヘリックスはタンパク質の基本構造の1つで、バネに似た右巻きらせんの形状をとる。両親媒性ヘリックスはその一形態で、らせんの側面のうち片側が疎水的、反対の側が親水的になっているαヘリックスを指す。MADS転写因子のKドメインは2つの両親媒性ヘリックスからなり、疎水的な側面同士の相互作用の可否により、特定のMADS転写因子間で多量体化が起きる。ファイロジェンは同様の構造をとることにより、特定のMADS転写因子を認識していると考えられる。