植物の低温ストレス耐性獲得におけるシグナル経路の通説を覆す発見
- 発表者
-
城所 聡(東京大学大学院農学生命科学研究科 応用生命化学専攻 助教)
金 俊植(理化学研究所 環境資源科学研究センター 研究員)
石川 朋奈(東京大学大学院農学生命科学研究科 応用生命化学専攻 修士課程;当時)
鈴木 孝征(中部大学大学院応用生物学研究科 応用生物学専攻 准教授)
篠崎 一雄(理化学研究所 環境資源科学研究センター センター長)
篠崎 和子(東京大学大学院農学生命科学研究科 応用生命化学専攻 教授)
発表のポイント
- 植物の低温ストレス耐性獲得における鍵転写因子DREB1Aをコードする遺伝子の発現は、これまで制御因子とされてきたICE1転写因子とは無関係であることが示されました。
- ICE1の変異体ice1-1でのDREB1A遺伝子の発現低下は、DREB1AプロモーターのDNAメチル化によるエピジェネティクスな制御によって起っていることを明らかにしました。
- 「ICE1転写因子がDREB1A遺伝子の低温に応答した発現を制御する」というこれまでの通説は誤りであり、再考する必要があることが示されました。
発表概要
東京大学と理化学研究所、中部大学の共同研究グループは、植物の低温ストレス耐性獲得における鍵転写因子DREB1Aの遺伝子発現が低下するice1-1変異体の原因が、DREB1Aプロモーターのメチル化であることを明らかにしました。植物は、季節の変動や異常気象などにより低温ストレスに晒されると、DREB1と呼ばれる転写因子をコードする遺伝子の発現を強く誘導することで、耐性獲得に働く遺伝子群の働きを活性化させ低温耐性を獲得します。これまで、ICE1という転写因子がDREB1の発現を促進する上流因子の1つであると考えられてきました。このICE1が変異したice1-1変異植物体ではDREB1Aの発現が抑制されており、その原因がICE1転写因子のアミノ酸変異であるとされてきました。今回、篠崎和子教授らの共同研究グループは、ice1-1変異体におけるDREB1Aの発現の低下はICE1のアミノ酸変異とは無関係であり、DREB1Aプロモーターが過剰なDNAメチル化を受けることで発現が抑制されていることを見出しました。この研究結果により、「ICE1が転写因子としてDREB1の発現を制御する」というこれまでの通説は誤りであり、再考する必要があることが示されました。
発表内容
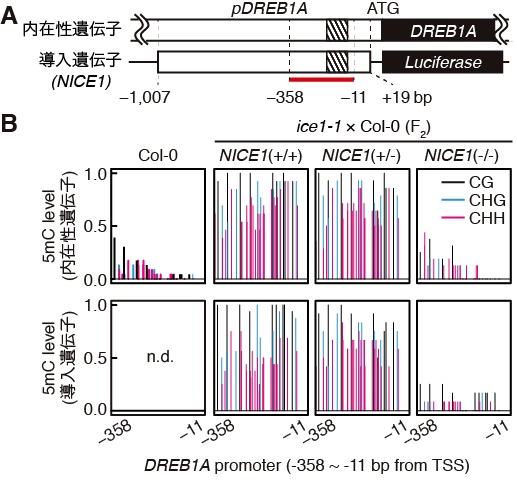
図1 NICE1 T-DNAによるDREB1AプロモーターのDNAメチル化。(A) 内在性のDREB1A遺伝子とNICE1 T-DNA上の導入遺伝子におけるDREB1Aプロモーターの模式図。網掛け部分は1ARを、赤線部分はメチル化レベルを測定した領域をそれぞれ示す。(B) DREB1AプロモーターDNAのメチル化レベル。ice1-1と野生型(Col-0)を掛け合わせた植物のF2世代のうち、NICE1 T-DNAを持つ個体((+/+)および(+/-))では、持たない個体(-/-)およびCol-0と比較してDREB1Aプロモーターのメチル化レベルが高い。
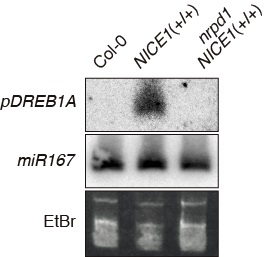
図2 NICE1 T-DNAによるDREB1Aプロモーター由来small RNAの産出。NICE1 T-DNAを持つ植物体ではDREB1Aプロモーター由来sRNAが検出された。RdDMの重要構成因子に変異を加えた植物体(NICE1(+/+) nrpd1)ではこのsRNAは検出されなくなった。
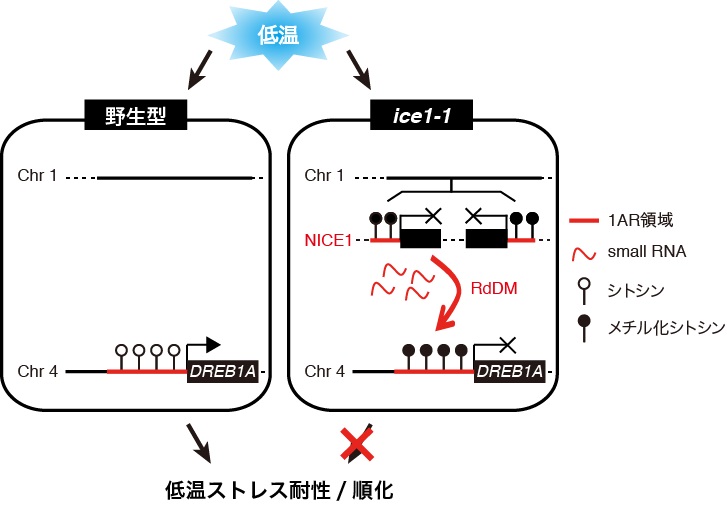
図3 ice1-1変異植物体におけるDREB1A発現抑制のモデル図。植物は低温ストレスに晒されるとDREB1A遺伝子の発現を誘導することでストレス耐性を獲得する。しかし、ice1-1変異植物体では、逆位のDREB1Aプロモーターを含むT-DNA座NICE1によって、DREB1Aプロモーター上に過剰なメチル化DNAが蓄積し、DREB1A遺伝子の発現が抑制された。
地球温暖化などの影響により異常気象が頻発しており、農業被害が深刻化しています。今後はさらに、急激な温度変化や旱ばつなどが頻発すると予測されています。このため、環境ストレスに高い耐性を示す作物の育種が重要となっています。植物は環境ストレスを受けると、様々な遺伝子の発現を誘導することにより耐性を獲得します。DREB1遺伝子は、植物の低温ストレスに応答した耐性の獲得機構において中心的な役割を持つ転写因子をコードしています。植物は、季節の変動や異常気象などによって低温ストレスに晒されると、DREB1遺伝子の発現を強く誘導することで、耐性獲得に働く多数の遺伝子群を強く働かせ低温耐性を獲得します。このため、DREB1遺伝子の発現を制御できれば、植物の低温に対する耐性を向上させることができると考えられます。
これまでに、本共同研究グループは、モデル植物のシロイヌナズナが持つ3つのDREB1遺伝子(DREB1A、DREB1B、DREB1C)のうち、DREB1BとDREB1C遺伝子の発現はCAMTAと呼ばれる転写因子によって急速な温度低下の時にのみ促進されることを見出しました。一方で、DREB1A遺伝子の発現はCAMTAによる制御を受けていませんでした。これまで、DREB1A遺伝子の発現を促進する主な上流因子はICE1と名付けられた転写因子であることが通説となっていました。その根拠として、最初に単離されたice1-1変異体と呼ばれる植物体ではDREB1A遺伝子の発現が抑制されており、その原因がICE1のアミノ酸変異であるとされていました。また、ICE1は葉の気孔分化にも関わっており、ice1-1変異体と同じ変異によって気孔分化に関わる下流遺伝子の発現を恒常的に促進することが知られていました。しかし、1つのアミノ酸変異が低温ストレス誘導性遺伝子の発現抑制と、気孔分化に関わる遺伝子の発現促進という逆の機能をもたらすメカニズムは不明のままでした。
今回、本共同研究グループは、シロイヌナズナのDREB1Aプロモーター中から低温ストレス誘導性の発現に関わる89塩基の領域(1AR)を同定しました。1ARにおける発現制御とICE1転写因子との関係を調べるために、1ARをつないだレポーター遺伝子を発現する植物体とice1-1変異体とを交配させた植物を作出しました。この植物のF2世代の個体を用いて、レポーター遺伝子とDREB1A遺伝子の発現量を解析したところ、両方の遺伝子の発現低下がICE1のアミノ酸変異とは独立して起こることを見出しました。一方で、気孔分化に関わる遺伝子の発現促進はICE1のアミノ酸変異に依存して起きていました。ICE1のアミノ酸変異とDREB1A遺伝子の発現低下との関係をさらに調べるために、別の研究グループから単離され、ice1-1変異体と同じアミノ酸変異を持つscrm-Dと名付けられた変異体の解析をおこないました。この変異体では、気孔分化に関わる遺伝子の発現促進が確認されましたが、DREB1A遺伝子の発現は低下していませんでした。したがって、ice1-1変異体内には、ICE1のアミノ酸変異とは異なるDREB1Aの発現抑制をもたらす原因があることがわかりました。さらに、ICE1の過剰発現または機能欠損させた植物体を用いてDREB1Aの発現を解析しましたが、ほとんど影響は見られませんでした。
そこで、ice1-1変異体内でDREB1Aの発現が抑制される本当の原因を探索するためにice1-1変異体の染色体DNA配列を再解読しました。その結果、DREB1Aの発現抑制が、DREB1A(第4染色体)やICE1(第3番染色体)の変異ではなく、第1染色体上にある1箇所のT-DNA座によって引き起こされることが示されました。このT-DNA座は複数のDREB1Aプロモーターを逆位に持っていました。このT-DNA座をNICE1と名付けました。染色体が異なる相同性配列による遺伝子制御としてエピジェネティクス制御の一種であるtransgene silencingの可能性を検証した結果、NICE1を持つ植物体ではDREB1Aプロモーター内にメチル化DNAが多く蓄積していました。NICE1を持った植物体に対して、DNAメチル化阻害剤で処理した場合やDNAメチル化経路RNA-directed DNA methylation(RdDM)の重要構成因子に変異を加えた場合には、DREB1AプロモーターのDNAメチル化が緩和され、DREB1Aの発現抑制が解除されました。さらに、NICE1を持つ植物では、RdDMの標的となるためのDREB1Aプロモーター配列を含む小分子RNA(small RNA; sRNA)が生産されていました。これらの研究結果から、ice1-1変異体におけるDREB1Aの発現抑制は、ICE1のアミノ酸変異ではなく、NICE1 T-DNAによるRdDMが誘発したDNAメチル化によって引き起こされたことが明らかになりました。また、これまでの「ICE1がDREB1Aの発現を制御する」という通説を再考する必要があると考えられました。今後は、1ARに結合する転写因子を同定することで、低温ストレスに応答してDREB1Aの発現を活性化する本当のメカニズムを明らかにできると考えられます。
発表雑誌
- 雑誌名
- 「The Plant Cell」(2月7日オンライン公開)
- 論文タイトル
- DREB1A/CBF3 is repressed by transgene-induced DNA methylation in the Arabidopsis ice1-1 mutant
- 著者
- Satoshi Kidokoro†, June-Sik Kim†, Tomona Ishikawa, Takamasa Suzuki, Kazuo Shinozaki, Kazuko Yamaguchi-Shinozaki*
- DOI番号
- 10.1105/tpc.19.00532
- 論文URL
- http://www.plantcell.org/content/early/2020/02/07/tpc.19.00532
問い合わせ先
東京大学大学院農学生命科学研究科 応用生命化学専攻 植物分子生理学研究室
助教 城所 聡(きどころ さとし)
Tel:03-5841-8111
Fax:03-5841-8009
E-mail:akido<アット>mail.ecc.u-tokyo.ac.jp <アット>を@に変えてください。
理化学研究所 環境資源科学研究センター 機能開発研究グループ
研究員 金 俊植 (きむ じゅんしく)
Tel:029-836-4359
Fax:029-836-2060
E-mail:june-sik.kim<アット>riken.jp <アット>を@に変えてください。
東京大学大学院農学生命科学研究科 応用生命化学専攻 植物分子生理学研究室
教授 篠崎 和子(しのざき かずこ)
Tel:03-5841-8137
Fax:03-5841-8009
E-mail:akys<アット>mail.ecc.u-tokyo.ac.jp <アット>を@に変えてください
用語解
- 注1 メチル化DNA
一般的にDNA中のシトシンの5番炭素にメチル基が付与された5-メチルシトシンを指します。真核生物の染色体DNA上でメチル化DNAが蓄積されることでヘテロクロマチン化を誘導し、トランスポソンなどの遺伝子の発現を抑えます。 - 注2 transgene silencing
組換え植物中で過剰に発現する外来遺伝子がDNAメチル化の標的となり、世代を経ることに外来遺伝子および相似した内在性遺伝子の発現が抑制される現象を示します。