メンデル遺伝の顕性を引き起こすDNAの「回文構造」を解明
- 発表者
-
藤井 壮太 (東京大学大学院農学生命科学研究科応用生命化学専攻 助教/科学技術振興機構(JST)さきがけ研究者)
下里 裕子(奈良先端科学技術大学院大学バイオサイエンス研究科 博士研究員:研究当時)
垣田 満(奈良先端科学技術大学院大学バイオサイエンス研究科 博士研究員:研究当時)
北西 高志(奈良先端科学技術大学院大学バイオサイエンス研究科 博士前期課程:研究当時)
岩野 恵(奈良先端科学技術大学院大学バイオサイエンス研究科 助教:研究当時)
高山 誠司(東京大学大学院農学生命科学研究科応用生命化学専攻 教授)
発表のポイント
- メンデルの顕性(優性)の法則はよく知られているが、その進化については未知の点が多い。本研究ではアブラナ科植物の自家不和合性の研究から、顕性が進化する分子メカニズムを新たに見出した。
- モデル植物のシロイヌナズナがSRKIRと呼ばれる特殊なDNA回文構造を進化の過程で獲得し、積極的に自家和合性になった可能性を見出した。
- 本研究では顕性形質の新しい分子進化パターンを発見した。DNAの回文構造、速やかな自然選択を受けるさまざまな顕性変異現象に関わる可能性が期待される。
発表概要
メンデルの顕性(優性)の法則は、多くの生き物に見られる普遍的な遺伝原理です。一方、このような関係がどのようにして出来上がるのかについて、分子的なメカニズムはあまりわかっていません。東京大学大学院農学生命科学研究科の高山誠司教授らの研究グループは、植物の自家不和合性と呼ばれる現象の研究から、顕性形質が進化する新しい分子メカニズムを見出しました。アブラナ科植物では雌しべの細胞膜にあるSRKと呼ばれる受容体が、自己の花粉表面をコーティングするタンパク質SP11を認識することで自己花粉拒絶反応が起きます。グループではモデル植物であるシロイヌナズナがどのようにして自家和合性を獲得したのかについて研究を行いました。研究の結果、多くのシロイヌナズナは逆位反復配列SRKIRと呼ばれる特殊なDNA回文構造を持ち、その構造によって顕性的にSRKの遺伝子発現を抑えていることが明らかになりました。逆位反復配列は顕性が進化する上でこれまで知られていなかった分子実態であり、その発見はさまざまな遺伝現象の説明へとつながると期待されます。本研究成果はオンライン誌Nature Communicationsに掲載されました。
本研究は、科学研究補助金基盤A「アブラナ科およびナス科植物の自家不和合性の分子機構解明」(研究代表者:高山 誠司、研究期間:2013年4月〜2016年3月)および基盤S「植物自家不和合性の分子機構と進化」(研究代表者:高山 誠司、研究期間:2016年5月〜2021年3月)の一環で行われました。
発表内容
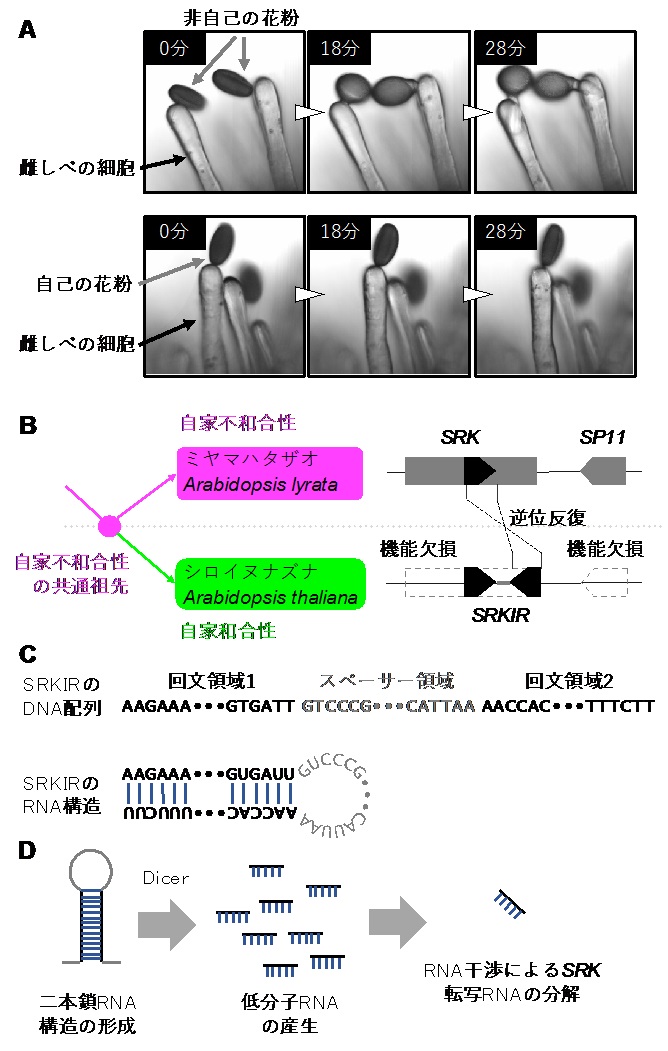
図A:アブラナ科の非自己の花粉(上パネル)と自己の花粉(下パネル)を雌しべの細胞に受粉したのちの経過写真。非自己を受粉した場合、花粉管が発芽し、雌しべに伸長する様子が見られる。自己の場合はそのような反応は見られない。B:シロイヌナズナでは近縁他種のミヤマハタザオのSP11—SRK遺伝子座に相当する領域を持つが、遺伝子機能は欠損している。また、多くの系統においてSRKに逆位反復が生じている。C:SRKIRは回文構造をとるため、RNAに転写されたのち、ヘアピン構造をとる。D:ヘアピン構造をとるRNAはDicerによって低分子RNAへとプロセッシングされる。その後、低分子RNAはSRKの伝令RNAに作用し分解する考えられる。
メンデルの顕性(優性)の法則は、多くの生き物に見られる普遍的な遺伝原理です。生物学の教科書などで、有名なエンドウマメの形質の分離比に関する研究が掲載されています。例えばマメの色が白い系統と、緑の系統を掛け合わせて得た交雑第二世代では、白と緑が3対1の比率で出現します。ふたつの形質のうちで多く現れる白は顕性、頻度が少ない緑は潜性と呼ばれます。現代では、着色の原因となるクロロフィルを分解する酵素が緑色の系統において壊れていることが現象の分子実態であるとわかっています(Sato et al PNAS 2007)。このように多くの場合、対立遺伝子の顕性・潜性関係は、遺伝子機能が壊れることで生み出されます。すなわち潜性が進化する方が大多数であると考えられてきました。しかし、稀に顕性が進化する例も報告されています。顕性をもたらす対立遺伝子は形質をあらわしやすいため、生物が急激な環境変化にすばやく適応するための鍵となる役割を果たしてきたと考えられます。それにもかかわらず、顕性が進化する分子メカニズムの多くはわかっていません。
研究グループは、植物の自家不和合性(注1)と呼ばれる現象を研究する過程で、新しい顕性形質の分子進化メカニズムを見出しました。半数以上の植物が持つ自家不和合性とは、自己と非自己を区別して受精するしくみです。多くの植物はこのしくみによって集団の個体間で積極的な遺伝子交換を行い、近親交配によって引き起こされる弱勢を防ぎながら、種の遺伝的な多様性を保っています。グループでは自家和合性でありアブラナ科のモデル植物でもあるシロイヌナズナ(Arabidopsis thaliana)に着目し研究を進めました。アブラナ科植物は雌しべの細胞膜にあるS—locus Receptor Kinase (SRK)と呼ばれる受容体が、自己の花粉表面をコーティングするタンパク質S—locus protein 11 (SP11)を認識することで自己拒絶反応が起きます(図A)。シロイヌナズナのすべての系統ではSRK遺伝子とSP11遺伝子のどちらかあるいは両方が壊れており自家不和合性を発揮できなくなっていることが知られています(図B)。一方、一部の系統では近縁他種ミヤマハタザオ(Arabidopsis lyrata)に由来する機能的なSP11とSRKを遺伝子組換えによって導入すると、自家不和合性を実験室内で再構成的に「復活」させることが可能です。しかし、その他の系統では両遺伝子を組み込んでも再構成ができないことが知られていました。研究グループでは、自家不和合性が再構成できない系統では壊れたSRK遺伝子の一部が逆位反復配列(inverted repeat)と呼ばれる構造を取っていることが原因であることを突き止めました。逆位反復配列とは、いわばDNAの回文構造です。このような回文構造はRNAに転写されたのち、鎖同士が相補的に結合することでヘアピンのような二本鎖構造(図C)を形成します。ヘアピンはDicerと呼ばれる酵素によって20から24塩基程度の細かいRNA分子(低分子RNA、注2)へと切断されます。この回文構造をSRKIR (SRK inverted repeat)と名付けて解析を進めたところ、SRKIRからは実際に20〜24塩基程度のRNAが大量に産生されていることが明らかとなりました。このような低分子RNAはRNA干渉(注3)というメカニズムなどによって相補的な転写RNAを分解することが知られています。本研究ではSRKIRに由来する低分子RNAが機能的なSRK遺伝子のRNA発現を抑えることで(図D)、自家不和合性を積極的に邪魔していることを明らかにしました。
さらにSRKIRはシロイヌナズナにおいて少なくとも2種類が平行に進化してきており、80%以上の系統がどちらかのタイプのSRKIRを持つことが推測されました。すなわちSRKIRの進化は偶然ではなく、必然的な選択であったと推察されます。花粉を運んでくれる生物(昆虫など)が激減する氷河期のような地球環境では、自家不和合性を抑えて自家和合性になる方が有利となると考えられています。シロイヌナズナという種は自家不和合性を顕性に抑制するSRKIRを獲得したことによって素早く種内に自家和合性を広めることができ、氷河期のように厳しい年代を生き抜くことができたのではないかと考えられました。
顕性が進化して集団の環境適応を可能にした有名な例として、蛾の一種であるオオシモフリエダシャク(Biston betularia)の工業暗化があります。この種の暗い体色は、産業革命の煤煙によって黒く汚れた樹木の上で保護色となり有利となるため、19世紀に急激に種内に広まったと言われています。この暗色は顕性形質ですが、cortexと呼ばれる遺伝子にトランスポゾン(注4)が挿入されることによりcortexのRNA発現が増加することで引き起こされることが報告されました(van’t Hof et al Nature 2016)。一方それとは異なり、本研究グループでは以前に低分子RNAが顕性の分子実態である例を報告しています(Tarutani et al Nature 2010; Yasuda et al Nature Plants 2016)。今回の報告はそれらのモデルを支持しつつ、新たに逆位反復配列と呼ばれるDNAの回文構造が顕性形質の原因メカニズムであることを突き止めました。逆位反復配列はDNA配列が突然変異によって重複と逆位により容易に生まれるため、さまざまな顕性変異の進化と速やかな拡散に関与してきた可能性が期待されます。
発表雑誌
- 雑誌名
- Nature Communications(3月16日)
- 論文タイトル
- Parallel evolution of dominant pistil-side self-incompatibility suppressors in Arabidopsis
- 著者
- Sota Fujii, Hiroko Shimosato-Asano, Mitsuru Kakita, Takashi Kitanishi, Megumi Iwano, Seiji Takayama*
- DOI番号
- 10.1038/s41467-020-15212-0
- 論文URL
- https://www.nature.com/articles/s41467-020-15212-0
問い合わせ先
東京大学大学院農学生命科学研究科 応用生命化学専攻 生物有機化学研究室
助教 藤井 壮太(ふじい そうた)
Tel:03-5841-5133
E-mail:a-fujii<アット>mail.ecc.u-tokyo.ac.jp <アット>を@に変えてください。
東京大学大学院農学生命科学研究科 応用生命化学専攻 生物有機化学研究室
教授 高山 誠司 (たかやま せいじ)
Tel:03-5841-5132
E-mail:a-taka<アット>mail.ecc.u-tokyo.ac.jp <アット>を@に変えてください。
用語解説
- 注1 自家不和合性
植物が自己の花粉と集団内の非自己の花粉を識別して、非自己のみと受精し子孫を作る性質。近親交配による有害な遺伝子の集積を回避し、集団内の遺伝的多様性を保つ効果があると考えられている。アブラナ科ではSP11という花粉タンパク質とSRKという雌しべタンパク質が自分自身のタイプのみと直接的に相互作用することで自己拒絶反応が起こる。 - 注2 低分子RNA
RNAは一本鎖として転写されるが、生体内では自己分子内で二本鎖RNAを形成するケースがしばしばみられる。このような二本鎖RNAはDicerと呼ばれるRNA分解酵素によって20-24塩基程度の小さなRNA断片へと切断される。 - 注3 RNA干渉
低分子RNAはRISC(RNA-induced silencing complex)と呼ばれるタンパク質複合体に取り込まれ、相同配列を持つ転写RNAの分解など、さまざまな生体反応を制御することが知られている。この効果はRNA干渉と呼ばれ、発見者は2006年にノーベル医学生理学賞を受賞した。 - 注4 トランスポゾン
ゲノム上を転移する特定のDNA配列であり、突然変異の原因となる。遺伝子領域にトランスポゾンが挿入されると機能の喪失あるいは変化、遺伝子発現の減少あるいは増加などさまざまな効果が引き起こされる可能性がある。