収斂進化で産み出された陸上植物のアレロパシー物質モミラクトン ~生合成経路の再構築でコケ類初の生合成遺伝子クラスターを同定~
- 発表者
- Lingfeng Mao(Institute of Crop Science, Zhejiang University博士課程大学院生:研究当時)
川出 洋(東京農工大学大学院農学研究院 教授)
樋口 俊哉(東京大学生物生産工学研究センター 修士課程大学院生:研究当時)
Meihong Chen(Institute of Crop Science, Zhejiang University修士課程大学院生:研究当時)
宮本 皓司(帝京大学理工学部 講師)
平田 佳樹(東京農工大学大学院農学研究院 修士課程大学院生:研究当時)
木村 穂乃花(東京農工大学大学院農学研究院 修士課程大学院生:研究当時)
宮崎 翔(東京農工大学大学院農学研究院 特任研究員:研究当時)
照屋 美優(東京大学生物生産工学研究センター 修士課程大学院生:研究当時)
藤原 薫(東京大学生物生産工学研究センター 修士課程大学院生:研究当時)
富田 啓介(東京大学生物生産工学研究センター 博士課程3年生)
山根 久和(帝京大学理工学部 教授:研究当時)
林 謙一郎(岡山理科大学理学部 教授)
野尻 秀昭(東京大学生物生産工学研究センター 教授)
Lei Jia(Institute of Crop Science, Zhejiang University 博士課程3年生)
Jie Qiu(Institute of Crop Science, Zhejiang University 助教)
Chuyu Ye(Institute of Crop Science, Zhejiang University 准教授)
Michael P. Timko(Department of Biology, University of Virginia 教授)
Longjiang Fan(Institute of Crop Science, Zhejiang University 教授)
岡田 憲典(東京大学生物生産工学研究センター 准教授)
発表のポイント
- 植物が生産する防御物質モミラクトン(注1)の生合成遺伝子群が、下等陸上植物の蘚類ハイゴケのゲノム上で遺伝子クラスターを形成することを明らかにしました。
- モミラクトン生合成遺伝子は高等植物のイネとイヌビエのゲノムでクラスターを形成して存 在しますが、本成果はコケ類を含む下等植物からの初めての生合成遺伝子クラスターの発見です。
- ハイゴケのモミラクトン生合成遺伝子クラスターを利用し、病気やストレスに強い作物を作 り出すことで、持続的農業の新展開につながる可能性が期待されます。
発表概要
盆栽に利用されるコケ植物の蘚類ハイゴケ(Calohypnum plumiforme、注2)は、周りの植物の成長を妨げるアレロパシー物質(注3)としてモミラクトンを生産します。今回、東京大学生物生産工学研究センターの岡田憲典准教授と東京農工大学の川出洋教授、浙江大学Longjiang Fan教授を中心とする研究グループは、ハイゴケのモミラクトン生合成遺伝子群がゲノム上で遺伝子クラスターを構成していることを明らかにしました。これまでに、イネと水田雑草のイヌビエがモミラクトン生合成遺伝子クラスターを持つことが同研究グループにより報告されていましたが、コケ類を含む下等植物での二次代謝化合物の生合成遺伝子クラスターの同定は、モミラクトンが初めての例となります。これらのモミラクトン生産植物では、生合成遺伝子の種類やゲノム上の配置が異なっていることから、陸上植物の進化の過程で独自に遺伝子をクラスター化し、収斂進化(注4)によりモミラクトン生合成能を獲得してきたと考えられます。さらに、ハイゴケ遺伝子をタバコに導入することでコケ由来の生合成経路をタバコ体内で再構築し、モミラクトンAまでの生産にも初めて成功し生合成の全容を解明しました。これらの成果は、陸上植物がどのようにモミラクトン生合成遺伝子クラスターを用いて生きるための競争力を獲得してきたのかを紐解く重要な手かがりとなるだけでなく、作物の生産における天然農薬としてモミラクトンを利用すると言った、環境調和型次世代農業への第一歩となることが期待されます。
発表内容
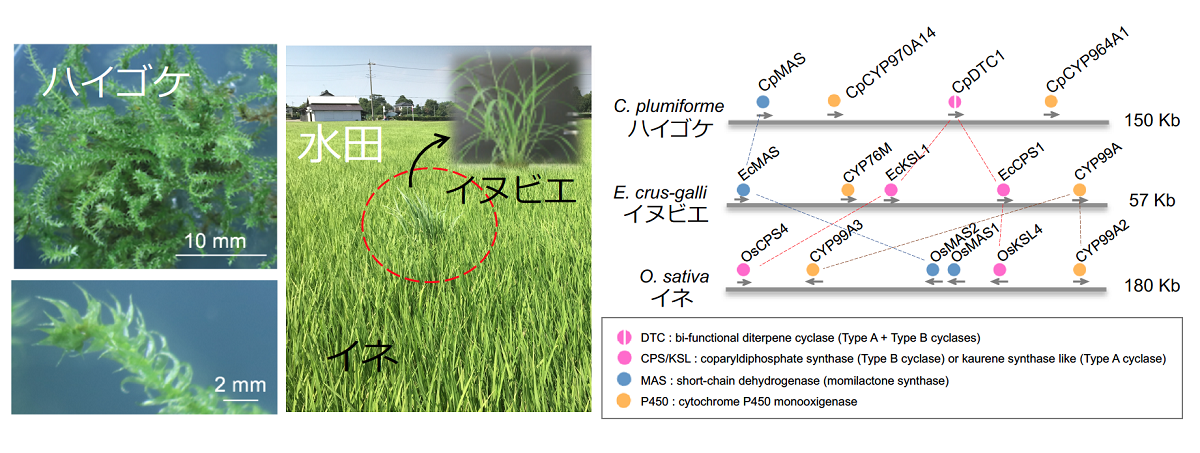
図1 モミラクトン生産植物における生合成遺伝子クラスター
下等植物の蘚類ハイゴケとイネおよび水田雑草のイヌビエといった、進化的に離れた3つの植物種がモミラクトンを生産することが知られている。【左図】
ハイゴケのモミラクトン生合成遺伝子クラスターは4つの遺伝子で構成されるが、イネやイヌビエのクラスター遺伝子とはゲノム上の配置が異なる。P450酸化酵素もそれぞれ異なるファミリーに属す酵素によりモミラクトンAが生産される。【右図】
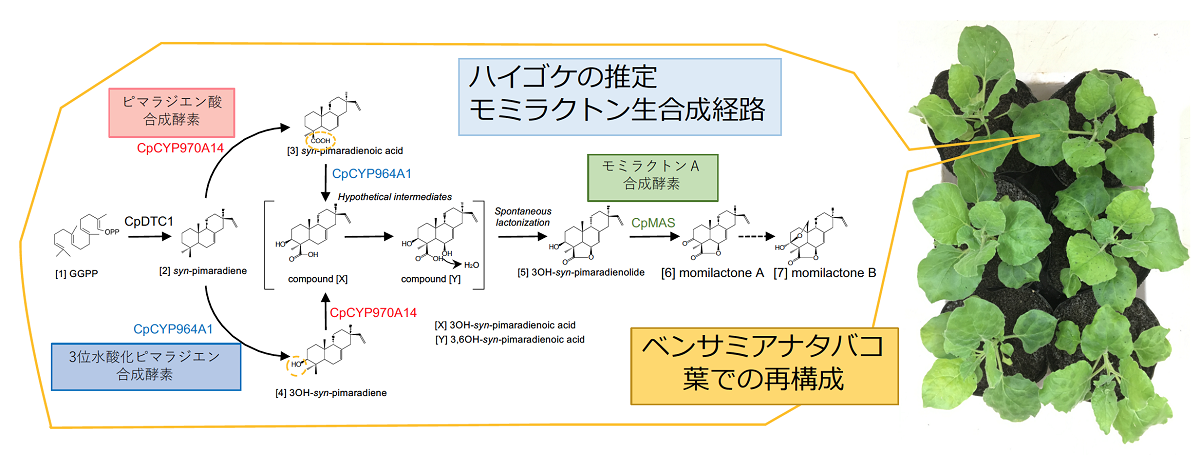
図2 ベンサミアナタバコでのモミラクトン生合成経路の再構成
ハイゴケのモミラクトン生合成遺伝子4種(CpDTC1, CpCYP970A14, CpCYP964A1, CpMAS)をアグロバクテリアによってタバコの葉に導入すると、モミラクトンAまでの異種生産が起こる。仮想中間体やモミラクトンB合成のステップを明らかにすることが今後の課題。
イネの抗菌性化合物として知られるジテルペン型ファイトアレキシン(注5)のモミラクトンは、水田雑草のイヌビエでも認められ、その生合成遺伝子が染色体上でクラスターを構成することが知られていました(図1)。モミラクトンは下等陸上植物である蘚類ハイゴケにおいてもアレロパシー物質としてその生産が報告されていましたが、モミラクトン生合成遺伝子としては、ゲラニルゲラニル二リン酸(GGPP、注6)からの初発の2段回環化反応により前駆体ピマラジエンの生産を担うCpDTC1が単離されているのみでした。進化的にかけ離れたイネとハイゴケにおいて、同一化合物であるモミラクトンがどのように生合成されるのかは、低分子の化学物質を利用した植物の抵抗性システムを考える上でとても興味深い謎であり、その解明には、まずハイゴケにおけるモミラクトン生合成経路の全貌解明が必要でした。また、既にモミラクトン生合成遺伝子クラスターの存在が明らかにされたイネとイヌビエでは、染色体上の遺伝子クラスターを構成する遺伝子の種類や配置が一致せず、その生合成経路の成り立ちに進化的なつながりがあるようには見えませんでした。そのため、第三のモミラクトン生産植物であるハイゴケのモミラクトン生合成関連遺伝子群が染色体上でクラスター化しているのかどうかは、とても重要で興味深い疑問でした。そこで我々は、ハイゴケのモミラクトン生合成遺伝子の探索と生合成経路の解明に着手しました。
まず始めに、ハイゴケの全ゲノム配列を決定し、遺伝子情報を網羅的に把握することでモミラクトン生合成遺伝子候補を探索しました。ハイゴケと同じ蘚類のコケの中でも、ヒメツリガネゴケはモデル植物としてよく用いられています。ハイゴケとヒメツリガネゴケのゲノム配列を比較した進化距離の解析では、この2つの蘚類のコケが分岐したのは約170万年前であり、これは単子葉植物と双子葉植物が分岐した時期とほぼ一致しました。この事は、蘚類のゲノム進化を考える上で、ハイゴケの情報がヒメツリガネゴケとは異なる新しい比較対象となる遺伝子情報として有用であることを意味します。先行研究で得られていたRNA-seqの情報も加味した遺伝子探索の結果、ピマラジエン合成酵素遺伝子CpDTC1以外に、モミラクトンA合成酵素ホモログのCpMASや数種のストレス誘導性シトクロムP450酸化酵素遺伝子をピックアップすることができました。さらに、ゲノム配列情報を基にこれらの遺伝子の染色体上の配置を調べて見ると、約150kbの領域にCpMAS, P450-1, CpDTC1, P450-2の順番で配置された遺伝子クラスターとして存在することがわかりました(図1)。そこで、これらの遺伝子が偶然隣り合わせに存在するのか、モミラクトン生合成遺伝子として機能しているのかを追究するため、酵素反応を調べました。大腸菌で発現させて精製酵素として得たHis-CpMASタンパク質については、モミラクトンAの前駆体であるピマラジエノライドを基質とした試験管内での反応においてモミラクトンA合成能が明らかになりました。2つのP450については、既知のP450アミノ酸配列との比較から約90%のidentityを保持するCpCYP970A14とCpCYP964A1と名付け、酵母とベンサミアナタバコを用いた解析を行いました。酵母のアッセイからは、CpCYP970A14がピマラジエンの19位炭素をカルボニルにまで酸化するピマラジエン酸合成酵素であること、また、CpCYP964A1が同じく3位の炭素を水酸化し3OH-ピマラジエンを与える活性を持つことがわかりました。さらに、これらの2種のP450遺伝子を、モミラクトン合成に必要なGGPP合成酵素遺伝子、さらにその上流のMEP経路遺伝子(注7)、CpDTC1、CpMASと共に同時にベンサミアナタバコに導入したところ、モミラクトンA生合成系路の再構成に成功し、タバコ内でモミラクトンAの生産を確認することができました(図2)。これらの結果から、ハイゴケのモミラクトンA生合成経路は4つの遺伝子で構成され、染色体上で遺伝子クラスターを形成していることが証明されました。この発見は、生合成遺伝子の機能が全容解明されていないイネ由来のモミラクトン生合成遺伝子クラスター研究に先駆けて生合成経路を明らかにした成果であり、さらにコケ類を含む下等植物において、初めての生合成遺伝子クラスターの存在を示す例となりました。
本成果により、現在までに発見されているモミラクトン生産植物の全てが、その生合成遺伝子を染色体上で遺伝子クラスターとして保持していることが示されました。同時に、それらのモミラクトン遺伝子クラスターにはシンテニーがなく、各植物の進化の過程で独立した収斂進化によって偶然にモミラクトンを獲得したものだと考えられます。あるいは、モミラクトンを合成するためには、遺伝子がクラスター化する必然性があるとも考えられます。今回、ベンサミアナタバコにおいて、モミラクトンを異種生産することにも成功しました。これは、モミラクトン生合成遺伝子がクラスターでなくても機能する証しでもあります。今後は、なぜモミラクトンAの生合成遺伝子がクラスター化しているのかを探求するため、クラスターを人為的に破壊したり、異なる染色体上に遺伝子を配置するなどして、クラスターの意味を追究したいと考えています。また、モミラクトンは抗菌化合物として有用なポテンシャルを持っているので、ハイゴケのモミラクトンA生合成遺伝子を利用して、新たなモミラクトン生産作物の作出にもチャレンジし、次世代農業での天然農薬としてのモミラクトン利用の可能性についても検討したいと考えています。
発表雑誌
- 雑誌名
- 米国科学アカデミー紀要 Proceedings of the National Academy of Sciences of the United States of America
- 論文タイトル
- Genomic evidence for convergent evolution of gene clusters for momilactone biosynthesis in land plants
- 著者
- Lingfeng Mao*, Hiroshi Kawaide*, Toshiya Higuchi*, Meihong Chen, Koji Miyamoto, Yoshiki Hirata, Honoka Kimura, Sho Miyazaki, Miyu Teruya, Kaoru Fujiwara, Keisuke Tomita, Hisakazu Yamane, Ken-ichiro Hayashi, Hideaki Nojiri, Lei Jia, Jie Qiu, Chuyu Ye, Michael P. Timko, Longjiang Fan**, and Kazunori Okada**#
*These authors contributed equally to this work.
** Correspondence
# Lead contact - DOI番号
- 10.1073/pnas.1914373117
- 論文URL
- http://www.pnas.org/cgi/doi/10.1073/pnas.1914373117
問い合わせ先
東京大学 生物生産工学研究センター 環境保全工学研究室
准教授 岡田 憲典(おかだ かずのり)
Tel:03-5841-3070
Faxl:03-5841-3070
E-mail:ukazokad<アット>mail.ecc.u-tokyo.ac.jp <アット>を@に変えてください。
研究室URL:http://park.itc.u-tokyo.ac.jp/biotec-res-ctr/kampo/
用語解説
- 注1 モミラクトン
イネの籾殻から発見された抗菌性化合物で、いもち病菌などのカビの生長を抑える活性をもつ天然の防御物質。植物に対しても成長抑制効果を持つことからイネに競争力を与えていると考えられている。幅広い生物活性から天然農薬として利用の可能性が期待されている。 - 注2 蘚類ハイゴケ
ハイゴケ科ハイゴケ属のコケで蘚類に分類される。ほふく性で這うようにふえることから「這い苔」と命名された。アジアで幅広く見られる苔で、日本では園芸用に流通しており、いたる所に生息する。 - 注3 アレロパシー
植物が他の植物の生長を抑える物質(アレロケミカル)を放出したり、あるいは動物や微生物を退けたり引き寄せたりする効果の総称。他感作用ともいわれる。 - 注4 収斂進化
祖先からの遺伝による共通性を持たない複数の異なるグループの生物が、独立した自然選択によってそれぞれの生物の利益のために働くような共通の形質(ここではモミラクトン合成能)を個別に獲得する現象。 - 注5 ジテルペン型ファイトアレキシン
ファイトアレキシンは、植物がさまざまなストレスに応じて生産する化合物。植物種毎にさまざまな種類の化合物が存在する。イネのファイトアレキシンは化学構造や生合成経路の違いから、ジテルペン系のモミラクトン類、ファイトカサン類とフラボノイド系のサクラネチンに分類される。今回研究の対象としたハイゴケのファイトアレキシンはジテルペン系のモミラクトン。 - 注6 ゲラニルゲラニル二リン酸
炭素数20のプレニル二リン酸で、ジテルペンの出発物質として重要な化合物。植物ホルモンのジベレリンや光合成色素のクロロフィルの生合成にも必須であり、主に葉緑体内で合成されたのち、環化などの反応を受けて、さまざまなジテルペン化合物へと形を変えていく。 - 注7 MEP経路遺伝子群
メチルエリスリトールリン酸(MEP)の合成に関わる7つの遺伝子。翻訳産物は葉緑体内に移行し、解糖系からの一次代謝産物であるピルビン酸とグリセルアルデヒド3リン酸から、イソプレノイドの基本骨格となるイソペンテニル二リン酸とその異性体ジメチルアリル二リン酸を生産する。葉緑体内のテルペノイド合成への基質供給を担うイソプレノイドの上流経路を構成する植物二次代謝産物の生産における源流として働く。