菜の花の自己と非自己を識別するしくみを解明 ~自家不和合性の自他識別機構を三次元構造から明らかに~
- 発表者
- 村瀬 浩司 (東京大学大学院農学生命科学研究科 応用生命化学専攻 特任准教授)
森脇 由隆 (東京大学大学院農学生命科学研究科 応用生命工学専攻 助教)
森 智行 (奈良先端科学技術大学院大学 先端科学技術研究科 助教:研究当時/ 現 東京大学定量生命科学研究所 助教)
三島 正規 (東京都立大学大学院理学研究科 化学専攻 准教授)
鈴木 剛 (大阪教育大学教育学部 教育協働学科 教授)
渡辺 正夫 (東北大学大学院生命科学研究科 分子化学生物学専攻 教授)
箱嶋 敏雄 (奈良先端科学技術大学院大学 理事・副学長)
高山 誠司 (東京大学大学院農学生命科学研究科 応用生命化学専攻 教授)
発表のポイント
- アブラナ科植物の自家不和合性(注1)において、花粉拒絶反応を誘導するリガンド(注2)・受容体の立体構造とそれらの相互作用について調べました。
- 自他の識別を行うリガンドと受容体のアミノ酸が特定され、自己と非自己を区別するしくみが理解できるようになりました。
- 自家不和合性の花粉拒絶反応を人為的にオンやオフにする人工リガンドの設計が可能になります。
発表概要
東京大学大学院農学生命科学研究科の村瀬浩司特任准教授・高山誠司教授らの研究グループは、アブラナ科植物の自家不和合性に関わるリガンド・受容体の複合体構造を決定し、コンピューターシミュレーション技術を用いて自家不和合性の自他識別機構(注3)を解明しました。
植物の多くは自己の花粉を拒絶して、非自己の花粉で受粉する自家不和合性と呼ばれる性質をもっています。アブラナ科植物の自家不和合性においては多数の遺伝子型をもつ受容体SRKとリガンドSP11が、それぞれ同じ遺伝子型のパートナーのみを認識することにより、花粉拒絶反応を誘導することが知られていました。しかしながら、これらの分子がどのようにして自己と非自己を識別しているのかについての詳細は不明でした。
本研究ではSRK-SP11複合体の3次元構造を、X線結晶構造解析(注4)の手法を用いて原子レベルの解像度で決定することに成功し、SRKがどのようにしてSP11を認識しているのかを明らかにしました。また、計11種類の遺伝子型について、SRKとSP11の相互作用をMDシミュレーション(注5)によって調べたところ、SRKは自己のSP11とのみ安定して相互作用できることがわかりました。
これらの結果は、植物の自他識別のしくみをタンパク質構造レベルで初めて明らかにした成果であり、受精の人為的制御を介した優良品種の開発・生産に繋がりうる成果であることが期待されます。
発表内容
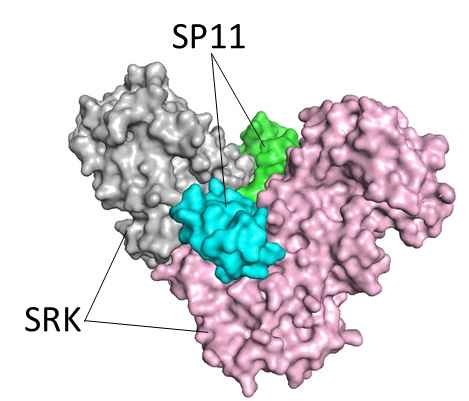
図1 S8型のSRK-SP11複合体
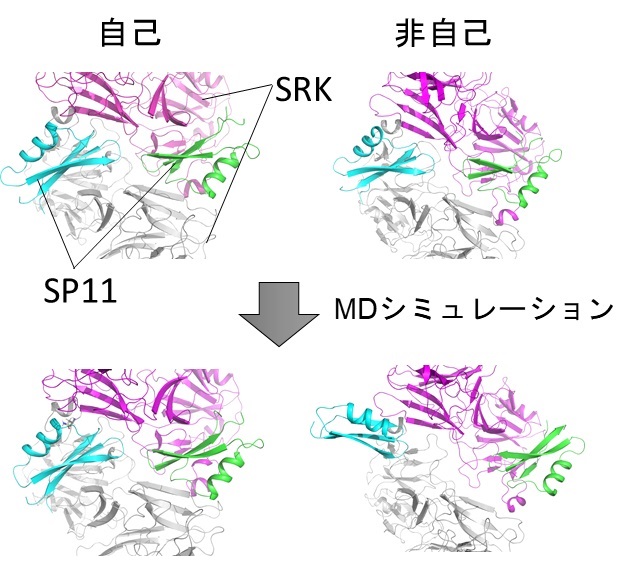
図2 MDシミュレーションによるSRK-SP11相互作用の観察
自己では複合体構造が維持されるのに対して、非自己ではSRKとSP11の間の親和性が低いためSP11が解離していく様子が観察された。
植物の花の多くは1つの花におしべとめしべがある両性花であり、自己の花粉で受精しやすい性質があります。多くの植物は種の遺伝的な多様性を確保するために、自己の花粉を拒絶して、非自己の花粉を受け入れる自家不和合性と呼ばれる性質を進化させてきました。アブラナ科植物の自家不和合性は多数のS遺伝子型(S1, S2, …, Sn)によって制御されており、めしべと花粉のもつS遺伝子型が一致すると、その花粉は拒絶されます。これまでの研究から自家不和合性の決定因子として、めしべ側の受容体SRKと花粉側のリガンドSP11が発見されています。また、めしべに存在するSRKは花粉から放出された同じ遺伝子型(自己)のSP11のみと結合して、花粉拒絶反応を引き起こすことが知られています。SRKとSP11はS遺伝子型によってアミノ酸配列が異なることから、このSRKとSP11による自他識別はアミノ酸配列の相違によって制御されていると説明されてきましたが、その詳細なしくみは不明でした。
本研究では菜の花(Brassica rapa)由来のS8型SRKとS8型SP11のタンパク質複合体を発現・精製して、0.2–0.3 mmのタンパク質結晶を作製しました。大型放射光施設(Spring-8、フォトンファクトリー)にて、この複合体結晶にX線を照射して回折データを収集し、2.6 Åの分解能(注6)でSRK-SP11複合体の立体構造を決定しました。SRKとSP11はそれぞれ2分子ずつの4量体を形成しており、V字型に配置したSRKの隙間にSP11が入り込んでSRKがお互いに離れないよう架橋していました(図1)。SRKにはS遺伝子型間で特にアミノ酸配列が異なる領域が3か所あり、これらの領域が主にSP11と相互作用していました。このS8型の構造をすでに構造が報告されているS9型のSRK-SP11複合体と比較したところ、タンパク質の全体的な骨格はよく似ているにもかかわらず、SRKとSP11が相互作用している領域は全く異なる形をしていることがわかりました。
次に、これらの構造を元に9つのS遺伝子型について新たにSRKとSP11の立体構造を再構築し、計11のS遺伝子型について自己と非自己の組み合わせで複合体を作製しました。次に、これらの複合体を用いてMDシミュレーションを行ったところ、自己の組み合わせでは安定な複合体を維持できるのに対し、非自己の組み合わせでは複合体を維持できずSP11がSRKから離れていく様子が観察されました(図2)。各SRK-SP11相互作用にかかる結合自由エネルギー(注7)を測定したところ、SRKとSP11のハプロタイプが自己の組み合わせのとき、最も安定であることがわかりました。また、これまでにSRKについてはSP11の認識に関わる領域が特定されているのに対して、SP11ではSRKの認識に関わる領域が不明でしたが、今回、アミノ酸レベルで結合自由エネルギーを解析することにより、SP11のSRK認識領域を同定することができました。さらに、SRKによるSP11の認識方式を分類すると、これらのS遺伝子型は3つのグループに分かれることも発見しました。
これら一連の解析結果から、SRKとSP11のS遺伝子型間を識別するアミノ酸の同定にも成功しました。例えばS32とS36型間ではSP11の62番目のアミノ酸が識別に関わっていると予測され、S36型SP11について62番目のアミノ酸をS32型のアミノ酸に置き換えるとS36型としてのSP11活性が消失することがわかりました。これらの結果はMDシミュレーションによる自他識別機構の予測が高い精度で行われていることを示しています。
アブラナ科植物の自家不和合性は雑種強勢を利用したF1ハイブリッド品種(注8)の作製に使われていますが、自家不和合性における自他識別のしくみはよくわかっていませんでした。今回の研究では、実際に行われている自他識別の瞬間をX線結晶構造解析とMDシミュレーションにより、高い精度で再現することに成功しました。また、MDシミュレーションによる観察から、様々なハプロタイプでSRKやSP11の自他識別に関わるアミノ酸の同定もできました。これらの情報を利用すれば、自家不和合性を人為的に誘導もしくは阻害を起こすようなSP11の設計が可能になり、優良品種の開発や生産に繋がることが期待されます。
この成果は英科学誌Natureのオンライン姉妹誌Nature Communicationsで2020年10月1日に掲載されました。
〈発表グループメンバー〉
村瀬 浩司 (東京大学大学院農学生命科学研究科 応用生命化学専攻 特任准教授)
森脇 由隆 (東京大学大学院農学生命科学研究科 応用生命工学専攻 助教)
森 智行 (奈良先端科学技術大学院大学 先端科学技術研究科 助教:研究当時/
現 東京大学定量生命科学研究所 助教)
劉 瀟 (奈良先端科学技術大学院大学 先端科学技術研究科 博士前期課程:
研究当時)
真坂 知帆 (奈良先端科学技術大学院大学 先端科学技術研究科 博士前期課程:
研究当時)
高田 美信 (東北大学大学院生命科学研究科 分子化学生物学専攻 技術専門職員)
前崎 綾子 (東京都立大学大学院理学研究科 化学専攻 研究員)
三島 正規 (東京都立大学大学院理学研究科 化学専攻 准教授)
藤井 壮太 (東京大学大学院農学生命科学研究科 応用生命化学専攻 准教授)
平野 良憲 (奈良先端科学技術大学院大学 先端科学技術研究科 助教:研究当時/
現 東京大学大学院薬学系研究科 助教)
河辺 善 (東京大学大学院農学生命科学研究科 応用生命化学専攻 博士前期課程:
研究当時)
永田 宏次 (東京大学大学院農学生命科学研究科 応用生命化学専攻 教授)
寺田 透 (東京大学大学院農学生命科学研究科 アグリバイオインフォマティクス教育研究ユニット 特任准教授:研究当時/
現 東京大学大学院農学生命科学研究科 応用生命工学専攻 准教授)
鈴木 剛 (大阪教育大学教育学部 教育協働学科 教授)
渡辺 正夫 (東北大学大学院生命科学研究科 分子化学生物学専攻 教授)
清水 謙多郎 (東京大学大学院農学生命科学研究科 応用生命工学専攻 教授)
箱嶋 敏雄 (奈良先端科学技術大学院大学 理事・副学長)
高山 誠司 (東京大学大学院農学生命科学研究科 応用生命化学専攻 教授)
発表雑誌
- 雑誌名
- Nature Communications(10月1日オンライン版)
- 論文タイトル
- Mechanism of self/nonself-discrimination in Brassica self-incompatibility
- 著者
- Kohji Murase, Yoshitaka Moriwaki*, Tomoyuki Mori, Xiao Liu, Chiho Masaka, Yoshinobu Takada, Ryoko Maesaki, Masaki Mishima, Sota Fujii, Yoshinori Hirano, Zen Kawabe, Koji Nagata, Tohru Terada, Go Suzuki, Masao Watanabe, Kentaro Shimizu, Toshio Hakoshima*, and Seiji Takayama*(*責任著者)
- DOI番号
- 10.1038/s41467-020-18698-w
- 論文URL
- https://www.nature.com/articles/s41467-020-18698-w
問い合わせ先
東京大学農学生命科学研究科 応用生命化学専攻 生物有機化学研究室
特任准教授 村瀬 浩司(むらせ こうじ)
Tel:03-5841-5134
E-mail:a-kmurase<アット>g.ecc.u-tokyo.ac.jp <アット>を@に変えてください。
東京大学農学生命科学研究科 応用生命工学専攻 生物情報工学研究室
助教 森脇 由隆(もりわき よしたか)
Tel: 03-5841-5448
E-mail:moriwaki<アット>g.ecc.u-tokyo.ac.jp <アット>を@に変えてください。
東京大学農学生命科学研究科 応用生命化学専攻 生物有機化学研究室
教授 高山 誠司(たかやま せいじ)
Tel:03-5841-5132
E-mail:a-taka<アット>g.ecc.u-tokyo.ac.jp <アット>を@に変えてください。
用語解説
- 注1 自家不和合性
花をつける植物では、めしべに花粉が付着すると花粉から伸びた花粉管がめしべの中に侵入して受精し、種子をつくる。多くの植物は自己の花粉を拒絶して、非自己の花粉で受精する自家不和合性と呼ばれるしくみを使って自殖を防いでいる。多くの場合、自家不和合性は複数の遺伝子型をもつS遺伝子座によって制御されており、花粉とめしべがもつS遺伝子型が一致すると花粉の拒絶反応が起こる。 - 注2 リガンド
細胞に存在する様々な受容体は多くの生体反応のスイッチとなっている。これらの受容体にはそのスイッチをオン/オフするための鍵穴があり、それぞれの受容体に特異的に結合する鍵分子がある。これらの鍵分子はリガンドと呼ばれ、受容体と結合することにより、受容体が制御する生体反応にオン/オフの切り替えを行っている。 - 注3 自他識別機構
自家不和合性を制御するS遺伝子座には花粉とめしべでそれぞれ自己と非自己を区別するための因子がある。アブラナ科植物ではS遺伝子座由来のめしべ側因子として受容体のSRKが、花粉側因子としてリガンドのSP11が発見されており、S遺伝子型によって異なるアミノ酸配列をもつことが知られている。SRKとSP11は同じS遺伝子型の相手とのみ結合して花粉拒絶反応を誘導することが知られているが、これらの分子がどのようにして自己と非自己を区別しているのかは不明であった。 - 注4 X線結晶構造解析
X線が結晶を通過する際に起こる回折現象を利用して、結晶中の分子構造を決定する方法。回折のパターンは結晶格子の形状と格子内の電子密度に依存するため、回折像を収集することにより、格子内の電子密度、すなわち分子構造を決定することができる。Spring-8などの大型放射光施設で高エネルギーの放射光を結晶に照射することにより、微小な結晶から分子の立体構造を決定できるようになった。 - 注5 MDシミュレーション
Molecular Dynamics(分子動力学)の頭文字をとってMDシミュレーションと呼ばれる。自然界で起こる物質の運動をコンピューター上に再現して、解析する手法の一つ。ここでは、タンパク質複合体の周囲に水分子やイオンを配置してタンパク質が実際に機能している場を再現した上で、分子を構成する1つ1つの原子について周囲の原子との間にかかる力を計算し、ニュートンの運動方程式を微小な時間刻みで数値的に解くことにより、分子の運動をシミュレーションした。 - 注6 分解能
物体をどの程度細かく見ることができるかの指標として用いられ、2つの点を視認できる最小の距離で表される。1 Å(オングストローム)は0.1ナノメートル(1ナノは百万分の1ミリメートル)。 - 注7 結合自由エネルギー
分子と分子が相互作用するのに伴う自由エネルギーの変化を意味する。結合自由エネルギーが負で絶対値が大きいほど高い親和性で結合できる。 - 注8 F1ハイブリッド品種
遺伝的な背景が異なる株を掛け合わせた子世代は雑種強勢と呼ばれる旺盛な生育を示すことが知られている。この雑種強勢を利用して作製されたF1種子はF1ハイブリッド品種と呼ばれる。F1種子の生産には自己の花粉で種子を作らないように、多くの場合、自家不和合性が利用されている。