システインが触媒の中心的役割を担う糖質分解酵素のメカニズムを解明
- 発表者
- Nicholas McGregor(ヨーク大学 博士研究員)
Joan Coines(バルセロナ大学 博士課程学生)
Valentina Borlandelli(ライデン大学 博士研究員)
味木 智子(東京大学大学院農学生命科学研究科応用生命工学専攻 大学院学生)
Marta Artola(ライデン大学 准教授)
Alba Nin-Hill(バルセロナ大学 博士課程学生)
Daniël Linzel(ライデン大学 大学院学生)
山田 千早(東京大学大学院農学生命科学研究科応用生命工学専攻 助教)
荒川 孝俊(東京大学大学院農学生命科学研究科応用生命工学専攻 助教)
石渡 明弘(国立研究開発法人理化学研究所 専任研究員)
伊藤 幸成(大阪大学理学研究科 特任教授)
Gijsbert van der Marel(ライデン大学 名誉教授)
Jeroen Codée(ライデン大学 教授)
伏信 進矢(東京大学大学院農学生命科学研究科応用生命工学専攻 教授)
Hermen Overkleeft(ライデン大学 教授)
Carme Rovira(バルセロナ大学 教授)
Gideon J. Davies(ヨーク大学 教授)
発表のポイント
- ビフィズス菌が植物の糖鎖を分解する酵素の反応メカニズムを詳細に明らかにしました。
- 触媒の中心的な役割を担うアミノ酸残基は「システイン」であり、これは糖鎖を加水分解する酵素としては初めての例です。
- 糖質分解酵素の応用範囲を広げ、新たなオリゴ糖合成手法の開発などにつながると期待されます。
発表概要
生物が糖鎖を分解する酵素は何百種類も知られており、食品分野やバイオ燃料生産などで幅広く用いられています。しかし、それらが糖鎖を加水分解する機構はどれもよく似通っていて、触媒に関わるアミノ酸残基もほぼ同じであることが知られていました。東京大学大学院農学生命科学研究科の味木智子大学院学生と伏信進矢教授らのグループは、ヨーク大学、バルセロナ大学、ライデン大学、および理化学研究所のグループと共同研究を行い、糖鎖の加水分解酵素としては初めての例として、システイン残基が触媒の活性中心として働く酵素の反応メカニズムを詳細に明らかにしました。この酵素の触媒残基に特異的に作用して結合する阻害剤を用いて反応中間体の立体構造を解明し、分子シミュレーション計算によりその中間体が外れる過程を解析しました。糖質分解酵素の新たな機構が分かったことにより、その応用範囲が広がることが期待されます。
発表内容
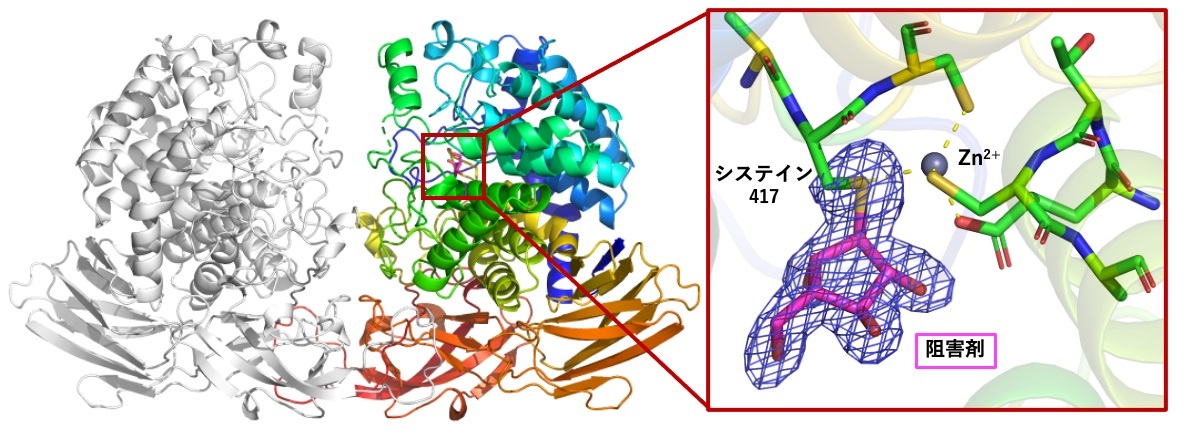
図1 HypBA1の立体構造(左)と、阻害剤が結合している活性部位(右)
阻害剤(マゼンタ)はHypBA1の417番目のシステイン残基に共有結合していた。触媒反応の中間体に類似したこの立体構造を元に、脱グリコシル化反応の分子シミュレーション計算が行われた。
糖質は生物の栄養源(澱粉など)、構造材料(セルロースなど)、細胞表面の情報伝達など、様々な機能を担う分子として利用されており、それらを分解する酵素(アミラーゼ、セルラーゼなど)も多数存在します。特に、糖鎖を加水分解により切断する酵素(糖質加水分解酵素)は非常に多くの種類が知られており、類似酵素をまとめたファミリー分類においても、現在160種類以上のファミリーが知られています(注1)。しかし、一方で、糖質加水分解酵素の触媒(化学反応)に関わる重要なアミノ酸残基(活性中心残基)は、ほとんどの場合「アスパラギン酸」と「グルタミン酸」であり、ほぼ共通の触媒機構を持つと考えられていました(注2)。
鹿児島大学の藤田清貴准教授らは、2011年に、ビフィズス菌から、食品に含まれる植物の細胞壁の糖鎖の一種である「β-アラビノオリゴ糖」を分解する酵素を発見しました。続いて、伏信教授らのグループは、藤田准教授および理化学研究所の石渡明弘研究員らと共同研究を行い、2014年に、ビフィズス菌のβ-アラビノオリゴ糖の分解酵素のひとつのHypBA1という酵素の立体構造を決定しました。その結果、HypBA1の活性中心残基が「システイン」であり、これが求核触媒残基として働く、という仮説を提唱しました(注3)。例えば、パパイヤなどのフルーツが持つタンパク質分解酵素(プロテアーゼ)では、求核触媒残基がシステインであることは古くから知られていました。一方で、糖質加水分解酵素でこのような例は知られていなかったため、この仮説を様々な実験手法で実証することが求められていました。特に、システインは求核触媒としての能力は高いものの、その求核性が強すぎるために、共有結合中間体が酵素から離れる「脱グリコシル化」がどのように起こるのかについて、よく分かっていませんでした(注4)。
今回、味木大学院学生と伏信教授らは、オランダ・ライデン大学のHermen Overkleeft教授らが開発・合成した新規な阻害剤「β-L-アラビノフラノシル型シクロフェリトール」を用いて、HypBA1と阻害剤が結合した立体構造を、X線結晶構造解析(注5)の手法を用いて明らかにしました(図1)。その結果、活性中心である417番目のシステインが阻害剤と共有結合を形成しており、このシステインが確かにHypBA1の求核触媒残基であることが分かりました。この立体構造は反応の中間体をよく反映していたため、この実験結果をもとに、スペイン・バルセロナ大学のCarme Rovira教授らは分子動力学(MD)計算と量子力学/分子力学(QM/MM)計算を行い(注6)、反応中間体の脱グリコシル化は、隣接する亜鉛イオン(Zn2+)の助けにより自由エネルギー変化の観点からも無理なく進行することを証明しました。英国・ヨーク大学のGideon J. Davies教授らは、HypBA1の類似酵素も同様の阻害剤と同様に反応することを示すなど、多くの生化学的実験データを提供して、4カ国のグループからなるこの共同研究を中心となって推進しました。
本研究によって、糖質加水分解酵素においてもシステインが求核触媒残基となりうることが証明されました。糖質加水分解酵素は、古くから澱粉工業などの食品産業で大量に用いられており、近年では、食品用のオリゴ糖や難消化性糖質の製造、バイオマスの変換、創薬分野など、様々なバイオ産業分野で利用されています。今後、糖質加水分解酵素を改変して利用する上で、本研究の知見をもとにして活性中心にシステインを利用することも可能になり、応用範囲がさらに広がると期待されます。
発表雑誌
- 雑誌名
- Angewandte Chemie
- 論文タイトル
- Cysteine Nucleophiles in Glycosidase Catalysis: Application of a Covalent β-L-Arabinofuranosidase Inhibitor
- 著者
- Nicholas G.S. McGregor, Joan Coines, Valentina Borlandelli, Satoko Amaki, Marta Artola, Alba Nin-Hill, Daniël Linzel, Chihaya Yamada, Takatoshi Arakawa, Akihiro Ishiwata, Yukishige Ito, Gijsbert A. van der Marel, Jeroen D.C. Codée, Shinya Fushinobu, Herman S. Overkleeft, Carme Rovira*, Gideon J. Davies*
- DOI番号
- 10.1002/anie.202013920
- 論文URL
- https://onlinelibrary.wiley.com/doi/full/10.1002/anie.202013920
問い合わせ先
東京大学大学院農学生命科学研究科 応用生命工学専攻 酵素学研究室
教授 伏信 進矢(ふしのぶ しんや)
Tel:03-5841-5151
Fax:03-5841-5151
研究室URL:http://enzyme13.bt.a.u-tokyo.ac.jp/
用語解説
- 注1 糖質加水分解酵素
糖と糖の間の結合を、水を用いて切断する酵素の総称であり、非常に種類が多い。2020年12月14日の時点で、CAZyというデータベースに記載されている糖質加水分解酵素の数は約88万6千種類であり、それらを類似酵素ごとにまとめたファミリーの数は161種類。酵素を活性の種類で分類した場合の糖質加水分解酵素(EC 3.2.1.-)の数は214。これらの数字は、他のタイプの酵素に比べても格段に多い。 - 注2 糖質加水分解酵素の活性中心残基
タンパク質は一般的に20種類のアミノ酸がつながってできている。通常の酵素はタンパク質であり、アミノ酸の側鎖(残基)が触媒反応に関わることが多い。糖質関連酵素の活性中心残基のほとんどは、側鎖にカルボキシ基を持つ、よく似た2種類のアミノ酸のいずれか(アスパラギン酸かグルタミン酸)である。詳細は以下のページで解説されている(英語サイト CAZypedia)
https://www.cazypedia.org/index.php/Glycoside_hydrolases - 注3 求核触媒残基
基質と反応して(求核攻撃を行い)共有結合を形成する触媒残基。アノマー保持型と呼ばれる一群の糖質加水分解酵素の中で、最も重要な触媒残基である。 - 注4 脱グリコシル化とシステイン残基
脱グリコシル化とは、基質と酵素の求核触媒残基が一旦共有結合で結びついた後に、水分子による求核攻撃を受けて、基質が酵素から離れる反応過程のこと。システイン残基は硫黄原子からなるチオール基という側鎖を持つため、求核攻撃の能力は高いが、脱グリコシル化がエネルギー的に不利なのではないかと疑問視されていた。 - 注5 X線結晶構造解析
酵素などのタンパク質の立体構造を明らかにするための最も一般的な解析方法の一つ。目的物質の結晶にX線を照射し、回折データを測定することにより、微細な三次元構造を知ることができる。 - 注6 分子動力学(MD)計算と量子力学/分子力学(QM/MM)計算
いずれも、多数の原子の動きや状態をコンピュータで数値計算して求める手法。MD計算は、原子の動きをニュートンの運動方程式にもとづいて求める。QM/MM計算は、量子力学的手法(QM)を分子力学法(MM)と組み合わせて、化学反応のシミュレーションや自由エネルギー計算を行う手法。