ソルガムの芒伸長はたった一つの顕性遺伝子で抑制できる
- 発表者
- 髙梨 秀樹 (東京大学 大学院農学生命科学研究科 生産・環境生物学専攻 助教)
堤 伸浩 (東京大学 大学院農学生命科学研究科 生産・環境生物学専攻 教授)
坂本 亘 (岡山大学 資源植物科学研究所 大気環境ストレスユニット 教授)
発表のポイント
- ソルガム (注1) において芒 (のげ) の伸長を強力に抑制する顕性 (注2) 芒抑制遺伝子、DOMINANT AWN INHIBITOR (DAI、注3) を同定しました。
- DAIは遺伝子重複 (注4) により誕生した新たな遺伝子であり、その芒抑制機能は新規に獲得したプロモーター (注5) によって引き起こされることを明らかにしました。
- ソルガムのDAIは異なる植物種であるイネの芒伸長も抑制できたことから、今後DAIを応用することでイネ科植物の無芒化に貢献できる可能性があります。
発表概要
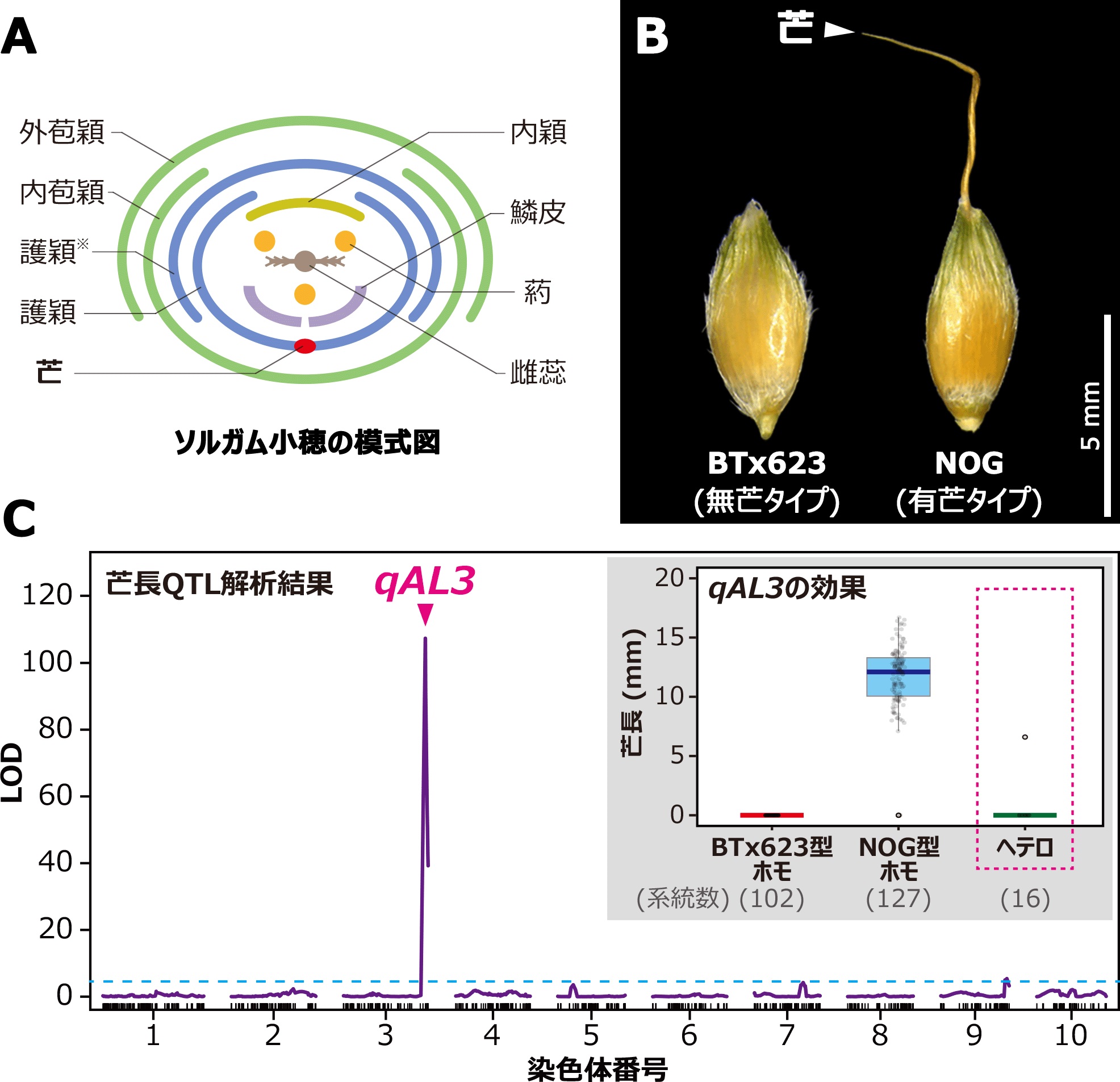
図1:ソルガム芒長に関するQTL解析結果
(A) ソルガム小穂の横断面模式図。各器官は外側から外苞穎、内苞穎、護穎 (※退化した第一小花の護穎)、護穎(芒はこの護穎から伸びる)、内穎、鱗皮、雄蕊 (葯)、雌蕊の順に着生している。
(B) 無芒タイプのBTx623 (アメリカで作出された系統) と有芒タイプのNOG (日本在来系統) の小穂の例。
(C) BTx623とNOGを二親としたRILsを用いた芒長QTL解析の結果。縦軸はLOD値 [基本的に値が大きいほど表現型 (今回の場合は芒長) に影響を与える遺伝子が近傍に存在する可能性が高い] を、横軸はソルガムの染色体上の位置を表し、水色の点線は有意水準を示す。第三染色体末端付近にLOD値116、寄与率91% (この集団における芒長の分散の91%がこのQTLによって説明される) という非常に効果の大きなQTL、qAL3が検出された。右挿入図: qAL3周辺の遺伝子型について、BTx623型ホモの個体群では芒長がゼロになり、NOG型ホモの個体群では芒が伸びることがわかる。興味深いことに、本ソルガム集団ではヘテロ型の個体群では芒長がゼロになるという、イネでは報告の無い「無芒型が顕性」であることが明らかになった。
(拡大画像↗)
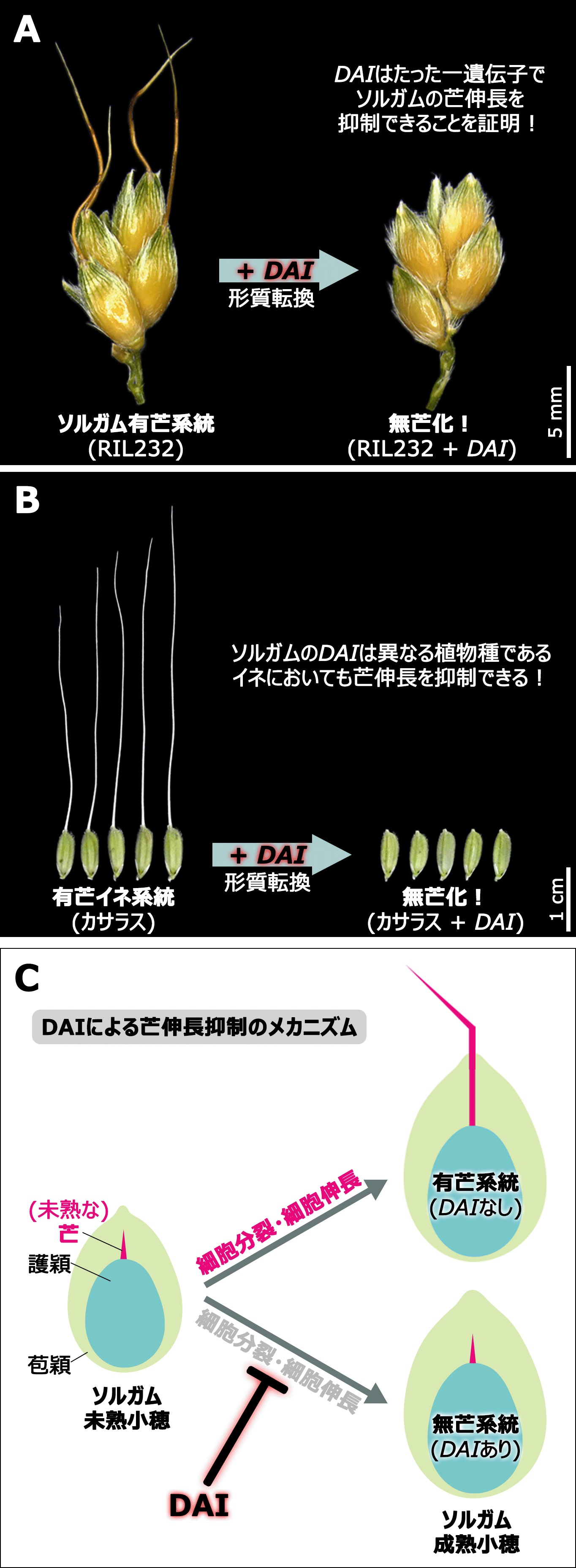
図2:顕性芒抑制遺伝子DOMINANT AWN INHIBITORの同定とその機能
(A) qAL3に含まれるある遺伝子を有芒ソルガムに形質転換したところ、芒が消失することが証明できたことから、この遺伝子をDOMINANT AWN INHIBITOR (DAI) と命名した。
(B) 興味深いことに、上記DAIを有芒イネ系統カサラスに形質転換したところ、ソルガムの場合と同様にその芒伸長を抑制できることがわかった。このことは、DAIによる芒伸長抑制機構は少なくともソルガムとイネの間で保存されていることを示している。
(C) ソルガム有芒系統および無芒系統における未熟小穂の経時的な解剖・観察結果から明らかになったDAIの機能。DAIは芒を構成する細胞に対して、その細胞分裂と細胞伸長の両方を阻害することで芒伸長を強力に抑制する機能をもつ。
(拡大画像↗)
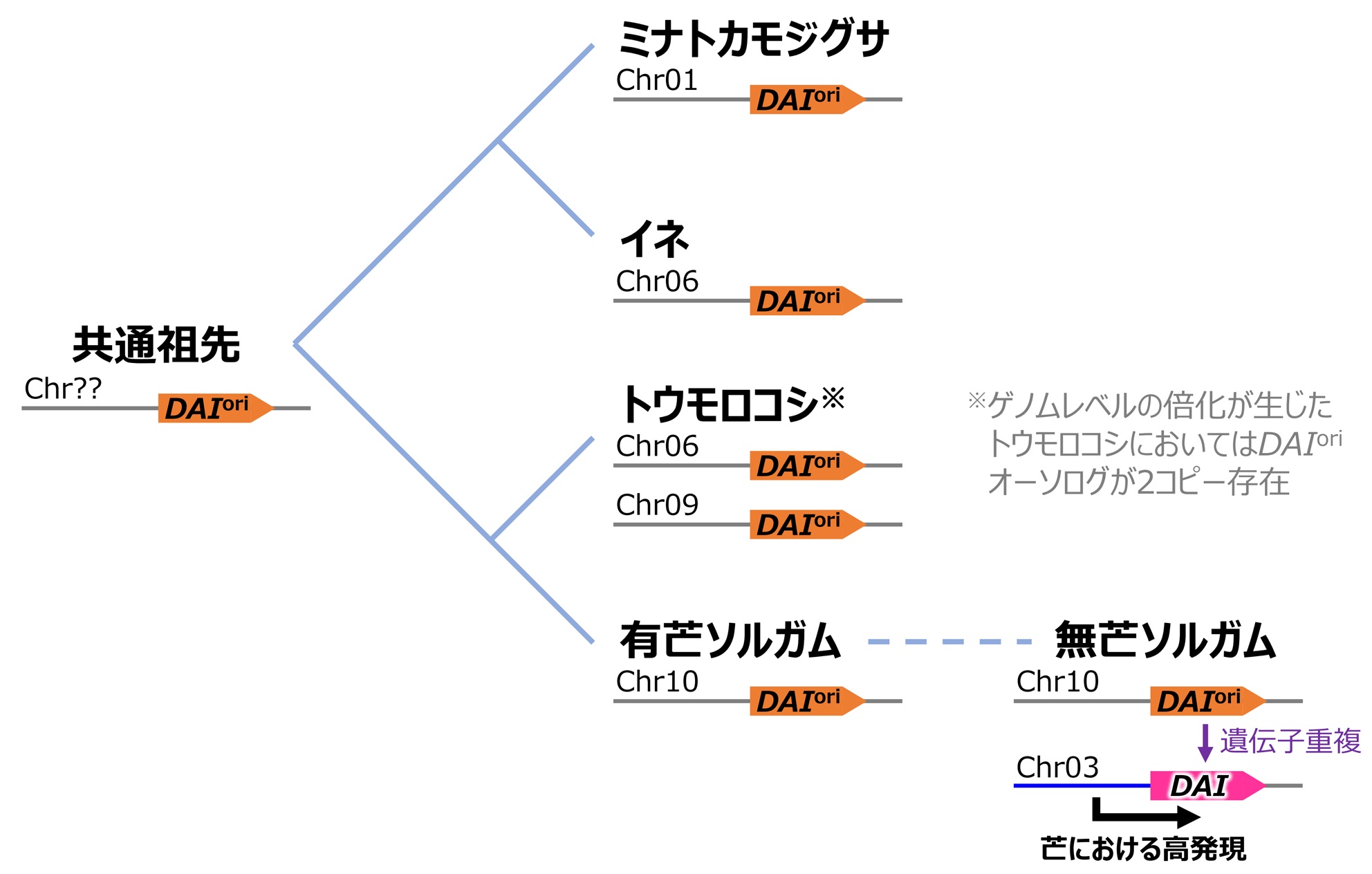
図3:遺伝子重複によるDAIの誕生
比較ゲノム解析 (シンテニー解析) の結果、ソルガムDAIは第十染色体に座上するDAIoriの遺伝子重複によって比較的最近誕生した遺伝子であることがわかった (他の近縁イネ科植物にはDAIオーソログが存在しない)。また、遺伝子発現解析により、この重複が生じた際にDAIoriのコピー (DAI) が挿入された部位 (第三染色体) の上流に「芒における特異的な高発現を誘導するプロモーター配列」が存在したことがDAIが新たな機能 (芒伸長抑制機能) を獲得するための鍵であったことが示唆された。(拡大画像↗)
イネ科植物の小穂 (花器官) から伸びる針状の構造である芒は、動物による食害に対する防御等に働く重要な構造であるものの、種子を収穫する人間にとっては邪魔な存在であったことから、例えばイネでは長い品種改良の歴史の中で芒が徐々に取り除かれてきたことがわかっています。イネとは異なり、ソルガムにおいてはこれまで芒を制御する遺伝子は同定されておらず、その制御に関する知見は非常に限られていました。
東京大学大学院農学生命科学研究科の髙梨秀樹助教、堤伸浩教授、および岡山大学資源植物科学研究所の坂本亘教授らの研究グループは、今回ソルガムにおいて非常に強い効果をもつ芒長関連遺伝子座を検出することに成功し、またこの遺伝子座に含まれる一つの遺伝子、DOMINANT AWN INHIBITOR (DAI) が顕性で芒伸長を抑制する責任遺伝子であることを明らかにしました。さらに、DAIは無芒ソルガム固有の遺伝子重複によって誕生した比較的新しい遺伝子であり、転移先に適切なプロモーター配列が存在したことが芒伸長の抑制という新たな機能を獲得するための鍵であったことが示されました。またDAIの機能が植物種間で保存されている可能性が示唆されたことから、本研究の成果はイネ科作物の芒に対して品種改良を行う際に非常に有用な情報となることが期待されます。
発表内容
イネ科植物の小穂から伸びる芒は、動物による食害に対する防御や種子の拡散等に働く重要な構造です。一方で、芒は人間にとっては収穫時に邪魔になる等のデメリットがあったことから、例えばイネではその栽培化 (注6) の過程で芒が徐々に取り除かれてきたことがわかっており、つまり芒は作物の栽培化の歴史を紐解く上でも興味深い形質であると言えます。
最も芒研究が進展しているイネにおいては、これまでに複数の「芒を伸ばす遺伝子」が同定されていますが、これらの遺伝子は基本的に1コピーでも存在すれば (ヘテロでも機能しうるため) 芒が伸長します (有芒型が顕性、注2)。したがって、イネの無芒化を達成するためには、栽培化の過程で複数の芒伸長遺伝子の機能を一つずつ完全に壊していく (機能喪失型の変異をホモ化する) 必要があったと考えられています。これらの遺伝子の機能解析を通じてイネの芒制御メカニズムについてはこれまでに多くのことがわかってきました。
一方で、世界五大穀物の一つであるソルガムにも無芒タイプや有芒タイプなど、芒の長さに関して大きな系統間差があるものの (図1AおよびB)、ソルガムにおいてはこれまで芒研究があまり進展しておらず、その制御に関する分子機構は明らかになっていませんでした。
東京大学、岡山大学などによる研究グループは、ソルガムにおける芒制御メカニズムを明らかにするため、組換え自殖系統 (注7) を用いて芒長に関するQTL解析 (注8) を実施しました。その結果、第三染色体上に非常に強い効果をもつ芒長関連遺伝子座qAL3 (Awn Length 3) を検出することに成功し、また驚いたことにqAL3の効果はイネでは報告の無い「無芒型が顕性」(つまり、この遺伝子が1コピーでも存在すれば芒が伸長しなくなる) であることも明らかになりました (図1C)。その後の解析で、qAL3に含まれる一つの遺伝子、DAIをソルガム有芒系統に形質転換することで無芒化が達成できたことから、このDAIが無芒化の責任遺伝子であることが証明されました (図2A)。更に、興味深いことに同様にDAIを導入した有芒イネ系統カサラスにおいても芒が綺麗に消失したことから (図2B)、DAIの機能は異なる植物種間でも共通して (保存されて) いる可能性が示唆されました。また、DAIは芒 (DAIをもつ「外見上無芒」のソルガム系統でも小穂内部には伸長が抑制された未成熟な芒がある) を構成する細胞に対して細胞分裂と細胞伸長の両方を阻害することで芒伸長を強力に抑制する機能をもつことが明らかになりました (図2C)。
続いて、ソルガム無芒系統のゲノム情報を精査したところ、DAIは第十染色体に存在する遺伝子 (DAIori) の重複によって新たに誕生した遺伝子であることが推測されました。これを検証するため、ミナトカモジグサ、イネ、トウモロコシ、およびソルガム (無芒および有芒) の四種のイネ科植物のゲノム情報を用いて比較解析を行ったところ、DAIoriとそのオーソログ (注9) は上記全てのゲノム中に存在したものの、DAIは無芒ソルガムのゲノムにしか存在しないことがわかりました。このことは、これらのイネ科植物の共通祖先種はDAIoriオーソログのみをもっており、DAIはソルガムと他のイネ科植物が種分化したのちに、無芒ソルガムの祖先型においてのみ生じた固有のDAIori重複によって誕生した比較的新しい遺伝子であることを示しています (図3)。
長い芒を有していたと考えられるイネ科祖先種の時点から存在していたDAIoriオーソログが芒伸長の抑制に機能しているとは考えにくく、そうなると「『もともと芒伸長の抑制とは無関係の機能をもっていたと考えられるDAIori』のコピーであるDAIが何故芒伸長を抑制する新たな機能を獲得できたのか」という疑問が湧きます。研究グループではこれを明らかにするため、DAIとDAIoriの両者をもつソルガム無芒系統の小穂を詳細に解剖し、定量的逆転写PCR法 (注10) を用いて小穂を構成する器官ごとに両遺伝子の発現量を定量しました。その結果、DAIはDAIoriがもともとほとんど発現していなかった芒で特異的に高発現していることがわかりました。この結果は、DAIoriのコピーが重複した際に、第三染色体の挿入部位の上流にたまたま芒での高発現を引き起こすプロモーター配列が存在し、この芒における新たな発現パターンの獲得がDAI特有の芒伸長抑制機能を生み出す鍵となったことを示唆しています (図3)。
DAIの応用展開に際して、最も大きなメリットはやはりこれが顕性の芒伸長抑制遺伝子であるという点です。例えば雑種強勢 (注11) を利用して種子収量を高めた無芒F1品種を作出することを考えた場合、これまでの (イネのような) 条件では、大変な手間をかけて「両方の」親系統について「複数の芒伸長遺伝子を全てホモで破壊」しておかなければF1は有芒になってしまいます。一方でDAIが適用可能である場合は、「どちらか一方」の親に「たった一遺伝子、DAIをホモで導入」しておくだけでF1は無芒となるため、F1の無芒化達成までのコストや時間が大幅に削減可能となります。本研究の成果は、イネ科植物の芒の制御機構や遺伝子重複を介した新規遺伝子の誕生に関する新たな知見をもたらすのみならず、今後イネ科作物に対して芒に焦点を当てた品種改良を行う際にも非常に有用な情報となることが期待されます。
【発表者一覧】
髙梨 秀樹 (東京大学 大学院農学生命科学研究科 生産・環境生物学専攻 助教)
鐘ケ江 弘美 (東京大学 大学院農学生命科学研究科 生産・環境生物学専攻 特任助教: 研究当時)
西村 明日香 (東京大学 大学院農学生命科学研究科 生産・環境生物学専攻 特任研究員)
山田 淳子 (東京大学 大学院農学生命科学研究科 生産・環境生物学専攻 学術専門職員)
石森 元幸 (東京大学 大学院農学生命科学研究科 生産・環境生物学専攻 特任研究員: 研究当時)
小林 正明 (明治大学 農学部 生命科学科 特任研究員: 研究当時)
矢野 健太郎 (明治大学 農学部 生命科学科 教授)
岩田 洋佳 (東京大学 大学院農学生命科学研究科 生産・環境生物学専攻 准教授)
堤 伸浩 (東京大学 大学院農学生命科学研究科 生産・環境生物学専攻 教授)
坂本 亘 (岡山大学 資源植物科学研究所 大気環境ストレスユニット 教授)
発表雑誌
- 雑誌名
- 「Plant and Cell Physiology」 (オンライン版: 5月27日)
- 論文タイトル
- DOMINANT AWN INHIBITOR encodes the ALOG protein originating from gene duplication and inhibits awn elongation by suppressing cell proliferation and elongation in sorghum
- 著者
- Hideki Takanashi, Hiromi Kajiya-Kanegae, Asuka Nishimura, Junko Yamada, Motoyuki Ishimori, Masaaki Kobayashi, Kentaro Yano, Hiroyoshi Iwata, Nobuhiro Tsutsumi*, and Wataru Sakamoto* (*責任著者)
- DOI番号
- 10.1093/pcp/pcac057
- 論文URL
- https://academic.oup.com/pcp/advance-article/doi/10.1093/pcp/pcac057/6595213?login=true
問い合わせ先
東京大学 大学院農学生命科学研究科 生産・環境生物学専攻 植物分子遺伝学研究室
助教 髙梨 秀樹 (たかなし ひでき)
E-mail: atakana<アット>g.ecc.u-tokyo.ac.jp <アット>を@に変えてください
Tel: 070-3284-5408
東京大学 大学院農学生命科学研究科 生産・環境生物学専攻 植物分子遺伝学研究室
教授 堤 伸浩 (つつみ のぶひろ)
E-mail: atsutsu<アット>g.ecc.u-tokyo.ac.jp <アット>を@に変えてください
Tel: 03-5841-5075
岡山大学 資源植物科学研究所 大気環境ストレスユニット
教授 坂本 亘 (さかもと わたる)
E-mail: saka<アット>okayama-u.ac.jp <アット>を@に変えてください
Tel: 086-434-1206
用語解説
- 注1 ソルガム (Sorghum bicolor)
熱帯アフリカ原産で、世界五大穀物の一つに数えられるイネ科の一年草 (別名タカキビ、コーリャン、モロコシなど)。光合成能力の高いC4植物で、他の主要穀物と比較して各種ストレス耐性が高いこともあり、世界中で広く栽培されている。食糧 (子実)、家畜飼料 (茎葉)、バイオエタノールの原料 (バイオマス作物) など幅広い用途に用いられている。 - 注2 顕性
異なる形質 (エンドウの種子の形が丸いかシワか などの特徴のこと) を示す両親 XとYを交配して得られた子供 (F1) が親Xの形質を示す場合、X側の形質を顕性 (優性) という。例えばイネのある遺伝子A [ここでは大文字Aは機能型の遺伝子を、小文字aは機能が壊れた (突然変異が生じたなどして) 遺伝子であることを意味する] が芒を伸ばす機能を持っているとする。あるイネ系統P1 (二倍体 = 体細胞ではゲノムが二セット存在) はこの遺伝子に関して機能型をホモ (AA) でもっており、別の系統P2は機能喪失型をホモ (aa) でもっていたとする。この場合、P1とP2を交配して得られたF1の遺伝子型はAa (ヘテロ) となる。遺伝子Aに関して、このF1は確かに機能喪失型遺伝子aをもっているものの、機能型遺伝子Aも1コピーもっているため、その機能型遺伝子が十分な働きを見せれば問題なく芒が伸ばせることになる。イネでこれまで知られていた芒関連遺伝子は全て「芒を伸ばす」タイプの遺伝子で、そういった遺伝子は基本的に1コピーでも存在すれば (ヘテロでも) 機能するため、イネでは有芒形質が顕性となる。今回同定したソルガムの芒関連遺伝子は「芒伸長を抑制する」タイプの遺伝子であり、1コピーでも存在すれば芒が消失することから、顕性芒抑制遺伝子DOMINANT AWN INHIBITORと命名した。 - 注3 DOMINANT AWN INHIBITOR (DAI)
DAIはALOGタンパク質の一つをコードする遺伝子であった。ALOGタンパク質とは、Arabidopsis LSH1 and Oryza G1 (ALOG) ドメインを持つ陸上植物特異的転写因子 (転写因子: 標的遺伝子の発現を誘導もしくは抑制するDNA結合性タンパク質) のこと。これまでの研究により、いくつかのALOGタンパク質は細胞分裂を抑制することで側生器官の発達を阻害することが報告されている。 - 注4 遺伝子重複
ある遺伝子を含むDNA領域がゲノム内で重複する現象のことを遺伝子重複と呼ぶ。大きなタイムスパンで見ると遺伝子重複は多くの生物で普遍的に見られる現象であり、例えばヒトでは全遺伝子の70%以上が遺伝子重複によって生じた遺伝子であると考えられている。 - 注5 プロモーター配列
一般的に転写 (DNA上の遺伝子配列からmRNAへ) 開始に関与する遺伝子の上流領域のことを指す。プロモーターはその遺伝子をいつ/どこで/どの程度転写させるかという時空間的な発現制御に対して大きな影響をもつ。 - 注6 栽培化
人為的な改良により野生植物が栽培植物 (人類にとって好ましい性質をもった植物) へと変化すること。有名な例としてイネの芒 (野生種: 有芒 → 栽培種: 無芒) や脱粒性 (種子が穂から自然に落ちる性質; 野生種: 脱粒性 → 栽培種: 非脱粒性) などが栽培化に関連した形質として知られている。 - 注7 組換え自殖系統 (Recombinant Inbred Lines: RILs)
異なる純系 (全ての遺伝子座がホモの個体) 同士を交配して得られた次世代個体 (雑種第一代: F1) を自殖すると、組換えによってさまざまなパターンで遺伝子型が分離したF2集団が得られる。RILsとは、各F2個体をスタートとしてそれぞれ自殖をくり返すことで得られる系統群を指す。十分な集団サイズで十分に自殖を繰り返した集団においては、さまざまな組換えパターンが保存されており、かつほぼ全ての遺伝子座がホモで固定されているため、遺伝学的な解析にとって非常に有用なツールとなる。 - 注8 QTL (Quantitative Trait Loci) 解析
ある集団における量的形質 (Quantitative Trait) の形質データとゲノムワイドなマーカー情報を用いて遺伝統計学的解析を行い、その形質を制御する責任遺伝子が座上しているであろうゲノム上の領域 (遺伝子座) を検出する手法。 - 注9 オーソログ
共通の祖先遺伝子から種分化に伴って派生した異種間の (同祖) 遺伝子同士の対応関係、あるいはそのような対応関係にある遺伝子群のことを指す。 - 注10 定量的逆転写PCR法
逆転写 (RNA → DNA) と定量的PCRを組み合わせ、特定の遺伝子の転写産物発現量を定量する方法 (ある遺伝子から転写されたmRNAが具体的に何コピー存在するかを数値化することが可能)。最近ではRNAウィルスであるコロナウィルスの検出に使われる手法としても有名。 - 注11 雑種強勢
ある特定の組み合せの両親を交雑した際に、雑種第一代 (F1) 個体が両親よりも優れた形質を示す現象のこと。この現象を利用した品種改良はF1ハイブリッド育種法とも呼ばれ、トウモロコシなどの穀物だけでなく、野菜・果樹・花卉などにも幅広く利用されている。