酸化ステロールの真の役割に迫る:コレステロールの水酸化反応によって引き起こされる脂質代謝変化を明らかに
- 発表者
- 齋藤 穂高(東京大学大学院農学生命科学研究科応用生命化学専攻 博士課程:研究当時)
立浦 和奏(東京大学大学院農学生命科学研究科応用生命化学専攻 修士課程)
西村 瑞希(東京大学大学院農学生命科学研究科応用生命化学専攻 修士課程)
清水 誠(東京大学大学院農学生命科学研究科応用生命化学専攻 特任准教授)
佐藤 隆一郎(東京大学大学院農学生命科学研究科応用生命化学専攻 特任教授)
山内 祥生(東京大学大学院農学生命科学研究科応用生命化学専攻 准教授)
発表のポイント
- コレステロール水酸化酵素によって細胞内で産生される酸化ステロールは、転写因子SREBP-2を選択的に阻害することで、コレステロール生合成を抑制することを明らかにしました。
- 複数のコレステロール水酸化酵素によって産生される酸化ステロールを網羅的かつ定量的に同定することに成功しました。
- 本研究より細胞内で産生される極微量の酸化ステロールの機能が明らかになったことで、酸化ステロールが関与する様々な疾患におけるその機能の理解が深まることが期待されます。
発表概要
酸化ステロール(注1)は、コレステロール恒常性を制御する重要なコレステロール代謝産物です。近年の研究より、酸化ステロールの中でも一部の水酸化コレステロールは、新型コロナウイルスをはじめとするウイルス感染や細菌感染を抑制する活性を有するほか、代謝疾患やがん、アルツハイマー病にも関与するなど、様々な疾患で多様な役割を果たしていることが示されています。しかし、酸化ステロールがどのようにこれらの疾患に関与するかは未だ十分に理解されていません。その原因として、細胞内で産生される酸化ステロールの機能に関する報告が少なく、その機能が十分に解明されていないこと、そして、生体内の酸化ステロール量が極微量(コレステロールの1/5,000〜1/10,000程度)で、その分析が困難であることが挙げられます。今回、東京大学大学院農学生命科学研究科の山内祥生准教授らのグループは、複数のコレステロール水酸化酵素(注2)を細胞に発現させ、コレステロールの狙った位置を水酸化させる(図1)ことで、細胞内で産生される様々な酸化ステロールの量や機能を詳細に解析し、細胞内で産生された酸化ステロールは転写因子SREBP-2(注3)を選択的に阻害することを見出しました。本研究成果は、細胞内で産生される複数の酸化ステロールの量や機能を明らかにした初めての報告であり、今後、酸化ステロールのさらなる機能解明や酸化ステロールが関与する病態の理解に貢献することが期待されます。
発表内容
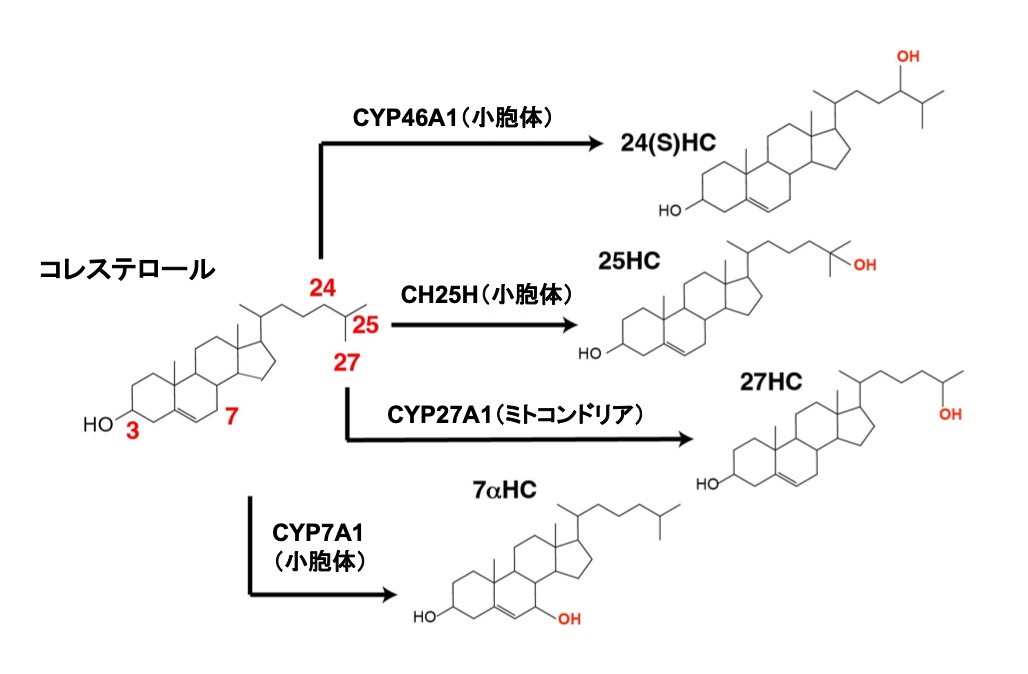
図1:コレステロール水酸化酵素によるコレステロール水酸化
コレステロール水酸化酵素CH25H、CYP27A1、CYP46A1、CYP7A1は、それぞれコレステロールの25位、27位、24位、7位を水酸化し、25-水酸化コレステロール(25-HC)、27-HC、24S-HC、7α-HCを産生する。CH25H、CYP46A1、CYP7A1は小胞体に、CYP27A1はミトコンドリアに発現する。
(拡大画像↗)
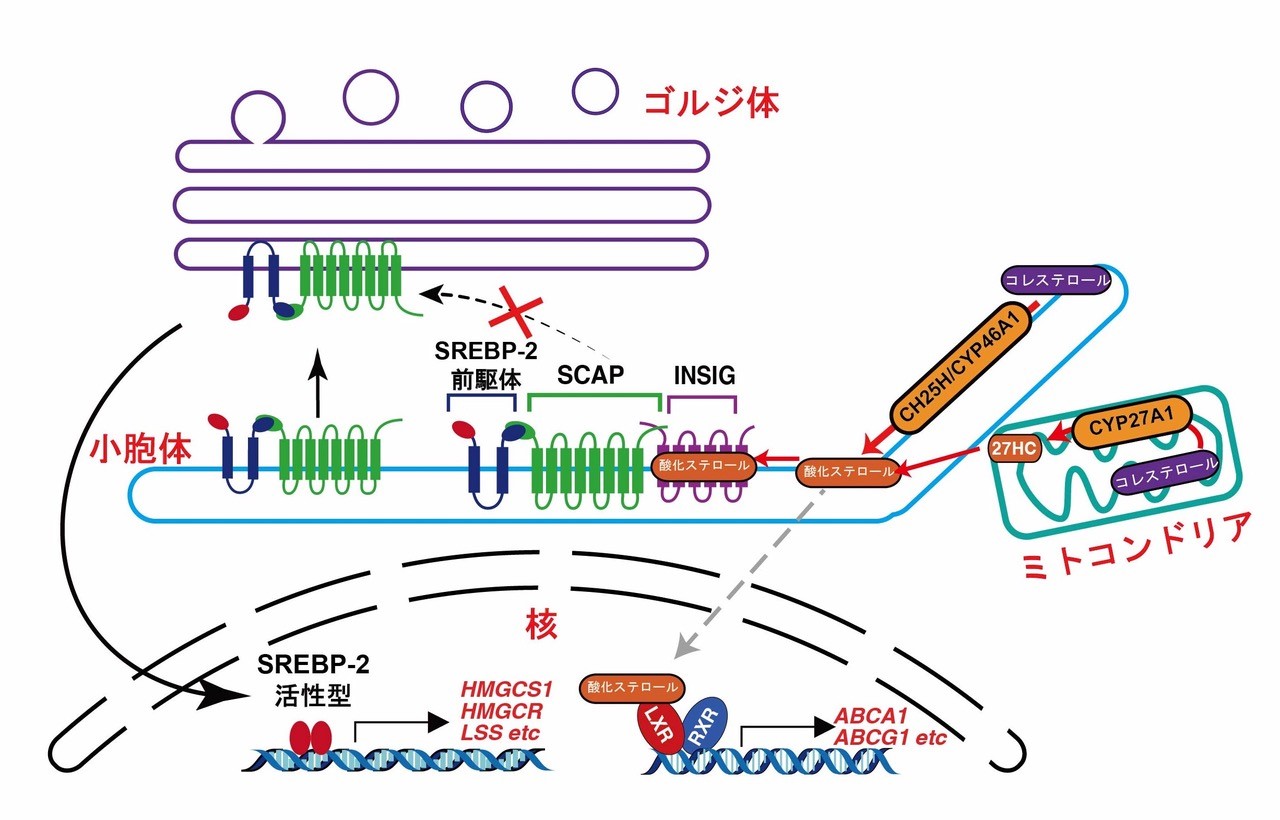
図2:細胞内で産生される酸化ステロールによるコレステロール恒常性制御の模式図
コレステロール水酸化酵素によって小胞体やミトコンドリアで産生された酸化ステロールは、INSIGと結合し、INSIGの分解を抑制する。その結果、SREBP-2の小胞体からゴルジ体への輸送は阻害され、SREBP-2活性化とそれに伴うコレステロール生合成関連遺伝子の発現が抑制される。一方、細胞内で産生された酸化ステロールはLXR標的遺伝子の発現には影響しない。
(拡大画像↗)
細胞コレステロールレベルは、コレステロールの生合成、細胞外からのコレステロール取り込み、そして、細胞外へのコレステロール放出によって厳密に調節されており、コレステロール恒常性の異常は動脈硬化症やがん、非アルコール性脂肪肝など様々な疾患で認められます。代表的なコレステロール代謝産物である酸化コレステロールは、コレステロールの生合成と取込みを包括的に制御する小胞体膜結合型転写因子SREBP-2を抑制するとともに、細胞外へのコレステロール放出を促進する核内受容体LXR(注4)を活性化することで、過剰なコレステロール蓄積を抑制すると考えられています。しかしながら、このような酸化ステロールの活性は、生理的濃度を超える過剰な酸化ステロールで細胞を処理することによって明らかにされた機能であり、細胞内で産生される酸化ステロールが同様の活性を有するかについて解析した報告はほとんどありません。
本研究では、細胞内でコレステロールを水酸化する主要な4種類の水酸化酵素(CH25H、CYP27A1、CYP46A1、CYP7A1)を細胞に発現させ、それらによって産生される25-水酸化コレステロール(25-HC)、27-HC、24S-HC、7α-HCがSREBP-2とLXRに及ぼす影響を詳細に解析しました(図1)。その結果、4種類のコレステロール水酸化酵素により産生された酸化ステロールの内、コレステロール側鎖が水酸化された酸化ステロール(25-HC、27-HC、24S-HC)は、SREBP-2活性を抑制する一方、LXR活性にはほとんど影響を及ぼさないことが示されました。また、このSREBP-2の不活性化は、SREBP-2活性を抑制する小胞体タンパク質であるINSIG(注5)の分解抑制によって引き起こされることが示されました。近年の研究より、細菌感染やウイルス感染によってCH25Hの発現が誘導され、25-HCが産生されること、さらに、25-HCが細菌感染やウイルス感染から細胞を守る活性を有していることが示されています。そこで、研究グループは、グラム陰性細菌の細胞壁成分であるリポ多糖でマクロファージ(注6)を刺激し、CH25Hの発現とそれに伴う25-HCの産生を誘導し、SREBP-2とLXR活性を評価しました。その結果、リポ多糖刺激に伴う25-HC産生によって、SREBP-2活性は抑制されましたが、LXR活性の変化は認められませんでした。さらに、研究グループは、質量分析装置を用いて、上記の4種類のコレステロール水酸化酵素やリポ多糖刺激によって産生される酸化ステロールを網羅的かつ定量的に解析し、これらの酵素の特異性を初めて明らかにしました。
酸化ステロールはSREBP-2活性を阻害することでコレステロール生合成を抑制するとともに、LXRを活性化することで細胞外へのコレステロール放出を促進するとこれまで考えられてきましたが、本研究より、細胞内で産生された酸化ステロールはSREBP-2活性を選択的に抑制し、LXR活性には影響しないことが示され(図2)、細胞内で産生される酸化ステロールのユニークな特性が明らかにされました。コレステロール水酸化酵素の発現は様々な病態で変動することが知られており、それに伴い産生される酸化ステロールの種類や量も変化することが予想されます。本研究成果は、酸化ステロールが関与する代謝疾患やウイルス・細菌感染、がんなどの病態のさらなる理解に貢献することが期待されるほか、酸化ステロールによって制御される特定の分子や分子経路を標的にした新しい病態制御の開発に応用されることも期待されます。
本研究は、革新的先端研究開発支援事業AMED-CREST(課題番号:20gm091008h, 21gm091008h)、日本学術振興会科学研究費助成事業(課題番号:19H02908, 22H02281, 20H00408)、日本学術振興会特別研究員奨励費(課題番号:20J10181)、アサヒグループ学術振興財団の支援により実施されました。
発表雑誌
- 雑誌名
- Journal of Biological Chemistry
- 論文タイトル
- Hydroxylation site-specific and production-dependent effects of endogenous oxysterols on cholesterol homeostasis: Implication for SREBP-2 and LXR
- 著者
- Hodaka Saito, Wakana Tachiura, Mizuki Nishimura, Makoto Shimizu, Ryuichiro Sato, Yoshio Yamauchi*(*:責任著者)
- DOI番号
- 10.1016/j.jbc.2022.102733
- 論文URL
- https://doi.org/10.1016/j.jbc.2022.102733
問い合わせ先
東京大学大学院農学生命科学研究科応用生命化学専攻 食品生化学研究室
准教授 山内 祥生(やまうち よしお)
Tel: 03-5841-5179
Fax: 03-5841-8029
Email: yoshio-yamauchi<アット>g.ecc.u-tokyo.ac.jp <アット>を@に変えてください。
用語解説
- 注1 酸化ステロール
コレステロールが酵素的・非酵素的に酸化されることで産生される。酸化ステロールうち、コレステロールが水酸化を受けたものを水酸化コレステロールという。生体内の主要な酸化ステロールは、25-水酸化コレステロール(25-HC)、27-HC、24(S)-HC、7α-HCであり、それらの量はコレステロールの約1/5,000〜1/10,000もしくはそれ以下である。これら4種類以外にも多種多様な酸化ステロールが存在しているが、その量はさらに少ない。 - 注2 コレステロール水酸化酵素
細胞内の小胞体やミトコンドリアに発現し、コレステロールの特定の部位を水酸化する。主要なコレステロール水酸化酵素は、本研究でも解析しているコレステロール25-水酸化酵素(CH25H)、シトクロームP450ファミリーのCYP27A1,CYP46A1,CYP7A1である。組織によって発現が異なり、CYP27A1は全身で発現するが、CYP46A1は脳特異的に、CYP7A1は肝臓特異的に発現する。CH25Hはリポ多糖やインターフェロンなどの刺激によって免疫細胞などで発現が誘導される。 - 注3 SREBP-2 (Sterol regulatory element-binding protein-2)
SREBP-2は、非常に珍しい膜結合型の転写因子で、小胞体膜に前駆体として発現する。細胞のコレステロールレベルが減少すると、小胞体からゴルジ体に輸送されて2段階の分解を受けることで転写活性ドメインが切り離され、活性化される。SREBP-2は、コレステロール生合成や取り込みに関連する遺伝子群の発現を包括的に制御している。 - 注4 LXR (Liver X Receptor)
LXRは、酸化ステロールをリガンドとする核内受容体の一つで、主に細胞外へのコレステロール放出に関わるトランスポーターの遺伝子発現を活性化する。 - 注5 INSIG (Insulin-induced gene)
INSIGは、コレステロール側鎖が水酸化コレステロールに高い親和性を示す小胞体膜タンパク質で、SREBP-2の活性化を抑制する働きをしている。酸化ステロールと結合することでINSIGの分解は抑制されて安定化するため、SREBP-2の活性化は阻害される。 - 注6 マクロファージ
体内に侵入したウイルスや細菌などの異物を貪食して消化することで、ウイルス・細菌感染を防ぐ機能を有する免疫細胞の一つ。免疫機能だけでなく、個体発生や組織の維持にも貢献している。