なぜ温暖な森林は生産量が大きいのか? ――樹木群集の炭素量分布が森林の生産量に貢献する――
- 発表者
- 東京大学大学院農学生命科学研究科・生圏システム学専攻
甲山 哲生(助教)
日浦 勉(教授)
ワーゲニンゲン大学
ダグラス シール(教授)
台湾・国立東華大学
孫 義方(教授)
国立研究開発法人森林研究・整備機構森林総合研究所
新山 馨(研究専門員)
群馬大学情報学部
西村 尚之(教授)
秋田県立大学生物資源科学部
星崎 和彦(教授)
鹿児島大学国際島嶼教育研究センター
鈴木 英治(特任教授)
台湾・国立嘉義大学
趙 偉村(助理教授)
台湾・国立林業試験所
伍 淑惠(助理研究員)
マレーシア森林研究所
ヌール ハージャル ザマ シャリ(主任研究員)
インドネシア国立研究革新庁生態学・民族生物学研究センター
ユニ セティヨ ラハユ(主任研究員)
北海道大学
甲山 隆司(名誉教授)
発表のポイント
- 東南アジア〜東アジアの60カ所の森林の継続調査データを用いて、樹木種レベルの年間炭素生産量が森林の炭素生産量をどのように構成しているかを解析しました。
- 樹木種の相対炭素生産量は、炭素量が小さい低木種ほど高い傾向を示し、この関係は熱帯から亜寒帯にかけて驚くほど共通していました。
- 熱帯林のように温暖な気候の森林では、種数の増加に伴って、低木種の比率が高くなるために、森林の生産量が大きくなることを、はじめて解明しました。
発表概要
東京大学大学院農学生命学研究科・生圏システム学専攻の甲山哲生助教をはじめとする北海道大学・ワーゲニンゲン大学・群馬大学など内外の共同研究者は、インドネシアやマレーシアの熱帯林から台湾や沖縄の亜熱帯林、そして鹿児島の暖温帯林から北海道の亜寒帯林に至る60の森林の継続調査のデータを用いた解析から、より温暖な森林ほど、相対的な(炭素量当たりの)年間炭素生産量が高い低木性樹種の比率が高くなることによって、同じ樹木炭素量を持つ森林の炭素生産量がより高くなることを明らかにしました。
従来の研究では、気温と樹木種多様性、そして森林生産の間の相関関係を示すに留まっていましたが、樹木種レベルの生産に注目した本研究では、これらの間の結びつきをはじめて機能的に解明しました。自然林の持続的管理では、炭素量が大きく、生産量も大きい高木種の保全が強調されてきましたが、本研究によって、低木種を含む樹木種多様性の保全が森林生産量の維持にとって重要であることが明らかにされました。
発表内容

図1:マレーシア・パソの熱帯多雨林(左)および北海道・雄阿寒岳の亜寒帯林(右) (拡大画像↗)
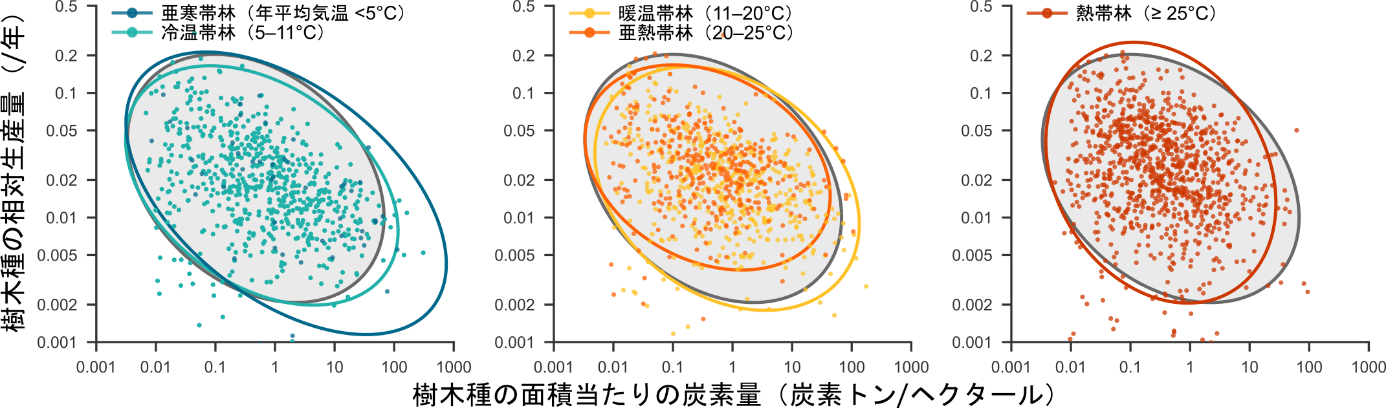
図2:樹木種レベルの相対生産量と面積当たりの炭素量の関係
各点は1森林調査地における1種の値を示す。楕円は2変量正規分布における95%信頼楕円を示し、色は調査地の年平均気温にもとづく森林の気候帯を示す。灰色の楕円はすべてのデータの95%確率楕円を示す。
(拡大画像↗)
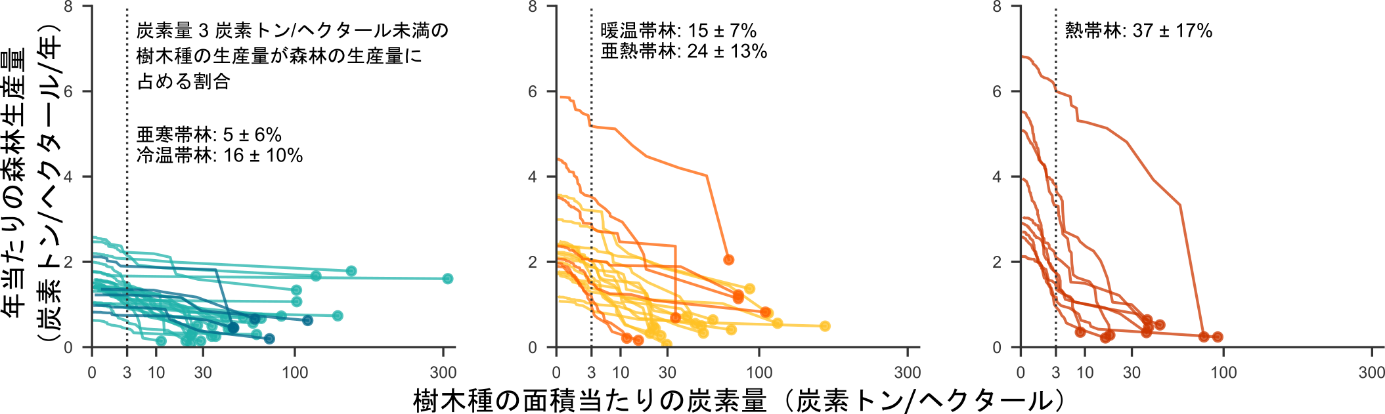
図3:森林を構成する樹木種の森林生産量への相対的な貢献度
炭素量の最小種から最大種の順に、森林生産量から種の生産量を差し引いていった減衰曲線。各線が1つの森林を示し、左端(縦軸との交点)が森林全体の生産量を、右端の円が炭素量最大の種の生産量を示す。色分けは図1と同じく森林の気候帯を示す。
(拡大画像↗)
〈研究の背景〉
森林の樹木は、陸上の生物炭素(注1)の生産と蓄積を担っており、地球上の炭素循環に中心的な役割を果たしています。とりわけ、熱帯多雨林は大きな森林の炭素生産量(注2)と高い樹木種の多様性を示します。
これまでの研究は、さまざまな森林の間で、樹木種多様性と炭素生産量の相関を調べるに留まり、これらの間の機能的な結びつきは未解明でした。また、森林の生産量の地域間比較にも手法的な問題がありました。樹木の繰り返し測定データに基づいて森林の純一次生産量(注3)を推定する従来の手法では、センサスの間に枯死した樹木の成長が無視されていました。さらに、森林を構成する個々の樹木種のターンオーバー速度(注4)の不均一性も考慮されてきませんでした。
このような推定手法上の問題を解消するために、著者らは樹木集団の生産量推定の新手法を提案しました。種多様性が高いマレーシアの熱帯多雨林にこの新手法を適用したところ、種レベルの相対的な(炭素量当たりの)炭素生産量が、炭素量の高い樹木種ほど減少することを見出しました。また、それぞれの樹木種について、その面積当たりの炭素量は、個体密度(単位面積当たりの本数)よりも種の最大樹高を反映することも示されました。こうした共存する樹木種間の機能分化に基づいて、森林を構成する低木種と高木種の炭素量の相対的な割合、すなわち「樹木群集構造」によって、森林生産量と種多様性そして気候との関連性が説明できるという仮説を立てました。一方で、別の可能性も考えられます。温暖な気候環境ほど、高い光合成速度や長い生育期間を反映して、おなじ炭素量を持つ樹木個体や樹木種でも生産量が高いことによって、温暖な森林ほど生産量が高くなっているのかもしれません。これらの可能性を検証すべく、本研究プロジェクトが立案・実施されました。
〈研究の内容〉
本研究では、インドネシアやマレーシアの熱帯林から台湾や沖縄の亜熱帯林、そして鹿児島の暖温帯林から北海道の亜寒帯林に至る60の森林の継続調査データを解析しました(図1)。これには環境省モニタリングサイト1000の森林調査サイトも40箇所含まれます。これらの地域はモンスーン気候の影響により一年を通して 森林が成立するのに十分な降水量があります。そのため、森林間の樹木種の多様性や生産量の違いは年平均気温によってよく予測することができ、年平均気温が高くなるほど多様性や生産量が高くなっています。
著者らが開発した新手法を用いて森林を構成するすべての樹木種(総計1,587種、2,604地域集団)の相対生産量を推定した結果、予測された通り、単位面積当たりの炭素量が大きい種ほど、相対生産量が小さい傾向が認められました。さらに、こうした種レベルの相対生産量と炭素量の反比例関係は、熱帯から亜寒帯に至る森林の間で驚くほど一致していました(図2)。
この結果は、森林の生産量が群集構造を反映しているという著者らの1つ目の仮説を強く支持していました。すなわち、より温暖な森林では、種数の増加に伴い、面積当たりの炭素量が小さくターンオーバー速度が速い低木種の比率が高くなるために、おなじ炭素量を持つ森林の生産量が高くなっていました(図3)。一方、温暖な環境ほど、樹木個体や樹木種が高い生産量を示すだろうという予想については、わずかにはその効果が認められたものの、森林の生産量の温度影響を説明するものではありませんでした。温暖な環境では光合成による有機物の稼ぎ分だけでなく、植物の呼吸による消費分も大きくなるため、稼ぎと消費の差引きである純一次生産量の温度依存性が明瞭でないと考えられます。
〈今後の展望〉
本研究の成果は、これまで表面的な相関解析によって推測されてきた森林の種多様性と生産量の関係に、個々の構成種の視点に基づいた機構的な解明を与えました。環境科学の重要課題である生物多様性と生態系機能の関係の解明に大きく貢献するものです。さらに、森林生態系の持続的管理という社会実装的課題(SDGsの目標15)に関しても、炭素量が大きく、生産量も高い高木種の保全が強調されてきましたが、本研究は、低木種を含む樹木種多様性と樹木群集構造の保全が森林の炭素生産の維持にとっても重要であることを指摘するものです。
研究助成
本研究は、日本学術振興会科学研究費助成事業(科研費)の学術変革領域研究(A)(課題番号:22H05715、21H05316)および基盤研究(B)(課題番号:18H02504、21H02248、21H02227)、基盤研究(C)(課題番号:19K06141)の支援により実施されました。
発表雑誌
- 雑誌名
- Nature Communications
- 論文タイトル
- Contribution of tree community structure to forest productivity across a thermal gradient in eastern Asia
- 著者
- Tetsuo I. Kohyama*, Douglas Sheil, I-Fang Sun, Kaoru Niiyama, Eizi Suzuki, Tsutom Hiura, Naoyuki Nishimura, Kazuhiko Hoshizaki, Shu-Hui Wu, Wei-Chun Chao, Zamah S. Nur Hajar, Joeni S. Rahajoe & Takashi S. Kohyama *責任著者
- DOI番号
- 10.1038/s41467-023-36671-1
- 論文URL
- https://www.nature.com/articles/s41467-023-36671-1
問い合わせ先
〈研究に関する問合せ〉
東京大学大学院農学生命科学研究科
助教 甲山哲生(こうやま てつお)
Tel:03-5841-8208
E-mail:kohyamat<アット>g.ecc.u-tokyo.ac.jp
〈報道に関する問合せ〉
東京大学大学院農学生命科学研究科・農学部 総務課 広報情報担当
TEL: 03-5841-8179 FAX:03-5841-5028
E-mail:koho.a<アット>g.ecc.u-tokyo.ac.jp
秋田県立大学 企画・広報本部 企画チーム
TEL: 018-872-1500 FAX:018-872-1672
E-mail:koho_akita<アット>akita-pu.ac.jp
北海道大学社会共創部広報課
TEL: 011-706-2610 FAX:011-706-2092
E-mail:jp-press<アット>general.hokudai.ac.jp
群馬大学情報学部 事務部広報担当
TEL: 027-220-7402 FAX: 027-220-7405
E-mail:si-web<アット>ml.gunma-u.ac.jp
<アット>を@に変えてください。
用語解説
- 注1 生物炭素
生物の生体を作る有機物に含まれる炭素(=現存量・biomass)。本記事中では、樹木の地上部を構成する生物炭素量を、単位面積当たりの質量(ヘクタール当たり炭素トン)で示す。 - 注2 炭素生産量
2回の樹木調査の間に、樹木個体の成長や、新たに加入した樹木個体によって増加した炭素量の増加分を、年当たりに換算した増加速度として表した値(著者らの新推定法が適用されている)。本記事中における生産量は、樹木の地上部純一次生産量(後述・注3)のうち、寿命の短い葉や繁殖器官などの生産量を差し引いた、樹木の成長と加入による生産量を指す(「地上部材生産量」とも呼ばれる)。 - 注3 純一次生産量
樹木が光合成によって固定した炭素生産の総量から、呼吸による消費分を差し引いた値を純一次生産量(純一次生産速度)という。 - 注4 ターンオーバー速度
時間経過に伴って、個体の成長・加入と死亡によって、生物炭素が入れ替わる速度。樹木の成長と加入によって増加した(材)生産量と、枯死によって損失された(=土壌に供給される)枯死量を足し合わせた値として表される。