葉緑体のゲノム編集 ~従来のゲノム編集酵素で置換が難しい塩基が置換可能に~
発表のポイント
- 葉緑体ゲノム上の特定の一塩基を別の種類に書き換える酵素 (標的一塩基置換酵素) を改良しました。
- 改変型の酵素は従来よりも酵素活性が向上し、これまで置換しにくいとされてきた塩基配列においても狙った一塩基を置換することができました。
- これまでよりも多くの塩基が置換対象の候補となり、また標的一塩基置換技術が葉緑体ゲノムの基礎研究に役立つことや、葉緑体ゲノムを利用した作物品種改良のための基盤技術になることが期待されます。
発表概要
東京大学大学院農学生命科学研究科の中里一星大学院生と有村慎一准教授らのグループは、モデル植物(注1)シロイヌナズナの葉緑体ゲノム約15万4千塩基対のうち狙った1塩基対のみを置換するゲノム編集酵素 (標的一塩基置換酵素) を改良しました。改変型の酵素は、従来の標的一塩基置換酵素が置換しにくい塩基を置換できることが示され、これまで考えられていたよりも多くの塩基が置換可能になります。標的一塩基置換酵素は、葉緑体ゲノムの基礎研究に役立つことや、葉緑体ゲノムの人為改変を利用した作物品種改良の基盤技術になることが期待されます。
発表内容
植物の細胞内で光合成を担う葉緑体は独自のゲノムを有しており、そこには光合成を行うために重要な遺伝子が存在します。これらの遺伝子を改変して植物のエネルギー生産効率を高めることができれば、作物の収量が向上する可能性があるため、人口増加により生じている食糧供給問題の解消や、地球環境への負担の少ないバイオ燃料の供給増加に貢献できる可能性があります。葉緑体のゲノム編集は、2021年に中里大学院生や有村准教授らのグループと韓国の別グループによって同時に世界で初めて達成、発表されました (https://www.a.u-tokyo.ac.jp/topics/topics_20210702-1.html)。これは、葉緑体移行シグナル配列を付加した標的一塩基置換酵素ptpTALECD(注2)を用いて、葉緑体ゲノム上の狙ったシトシン (C、注3) をチミン (T、注3) に置換する手法で、細胞当たり数百から数千個ある葉緑体ゲノムの全てで標的塩基を置換することができます (図1)。ptpTALECDはTやAの直後に位置するCを置換できることが分かっていましたが、グアニン (G) やCの直後のCを置換するのは酵素の性質上難しいとされており、置換できるかどうかはきちんと確認されていませんでした。本研究では、ptpTALECDに加えて、改良型のptpTALECD [ptpTALECD_v2、高活性型の塩基置換ドメイン(Mok et al., 2022, Nature Biotechnology, 40, 1378-1387)を有する] を用いて、酵素活性が実際に向上するかどうか、並びにこれまで置換されたという報告がなかった、GやCの直後のCを置換できるかどうかを検証しました。
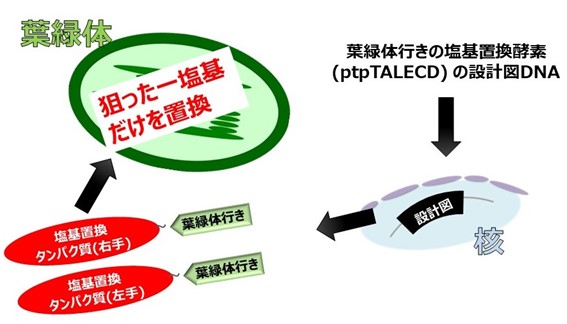
図1 ptpTALECDを用いた塩基置換の流れ
まず、塩基置換タンパク質ptpTALECDの設計図となるDNAをシロイヌナズナの核ゲノムに導入した。細胞内で作られた一対のptpTALECDが、葉緑体移行シグナル配列の情報に従って葉緑体の中に輸送される。最終的に、シロイヌナズナの葉緑体ゲノム約15万4千塩基対のうち標的の一塩基対のみを、細胞当たり数百から数千個あるゲノムコピーの全てで置換することに成功した [Nakazato et al., Nature Plants, 7, 906-913 (2021)]。
本研究ではまず、ptpTALECDまたはptpTALECD_v2の設計図となるDNAを、モデル植物シロイヌナズナの核ゲノムに導入しました。植物の細胞内で作られたptpTALECD (またはptpTALECD_v2) タンパク質は、自身が持つ葉緑体移行シグナルの配列情報に従って葉緑体の中へ運ばれると、二つのTALEドメイン (注2) が葉緑体ゲノム上の標的DNA配列に結合します。すると、二つのTALEドメインが結合した配列の間の、14~16塩基対の配列にある特定のCがTに変換されます。ptpTALECD_v2はptpTALECDと同様に、TやAの直後に位置するCを、一細胞に多数個存在する葉緑体ゲノムの全コピーで置換できることが分かりました。本研究で標的とした配列のうちいくつかでは、ptpTALECDはGやCの直後のCを一細胞に多数個存在する葉緑体ゲノムの一部コピーで置換しましたが、ptpTALECD_v2はこれらのCを一細胞に多数個存在する葉緑体ゲノムの全コピーで置換しました (図2)。ただし、標的配列によっては、ptpTALECDもGやCの直後のCを一細胞に多数個存在する葉緑体ゲノムの全コピーで置換することができました (図3)。また、ptpTALECD_v2はptpTALECDと比べて塩基置換活性が高いことも明らかになりました。
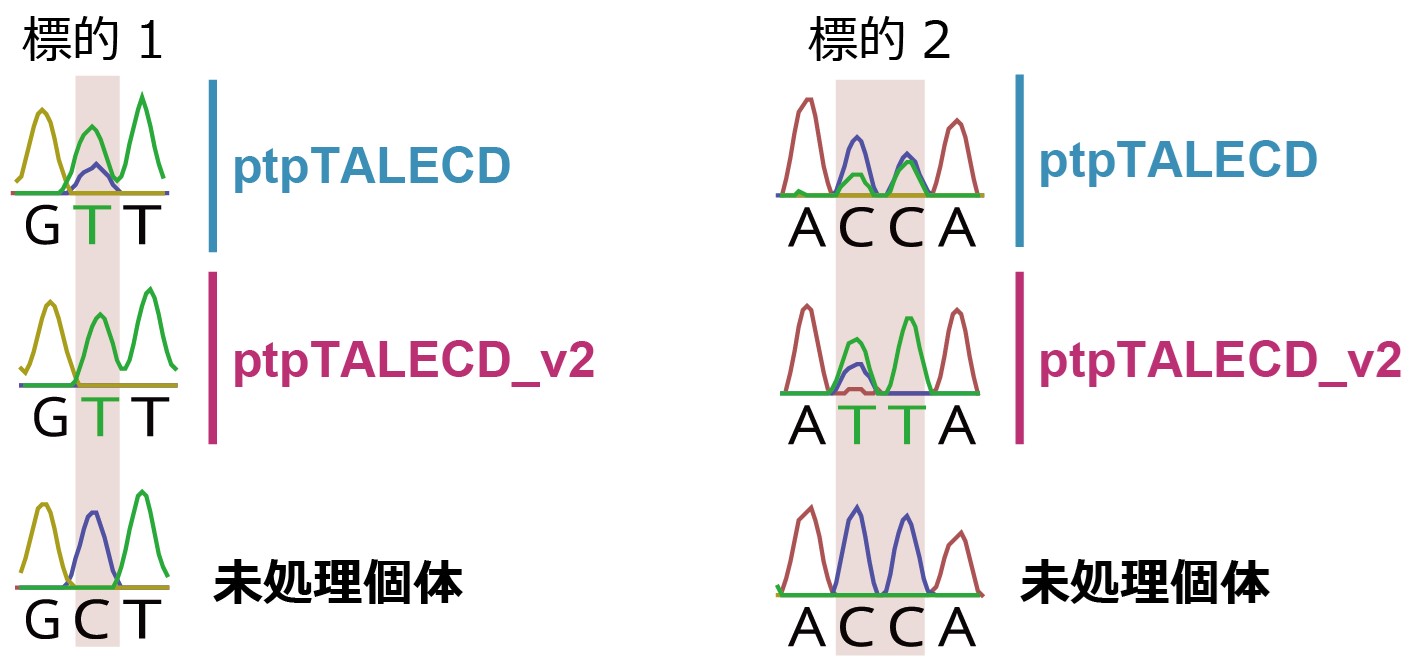
図2 ptpTALECDとptpTALECD_v2による、GやCの直後のCの置換
ptpTALECD導入個体(上)では、難標的とされていたGやCの直後のCが細胞内の葉緑体ゲノムコピーの一部でTに置換されたが、ptpTALECD_v2導入個体(中)では細胞内の葉緑体ゲノムコピーの全てでTに置換された。
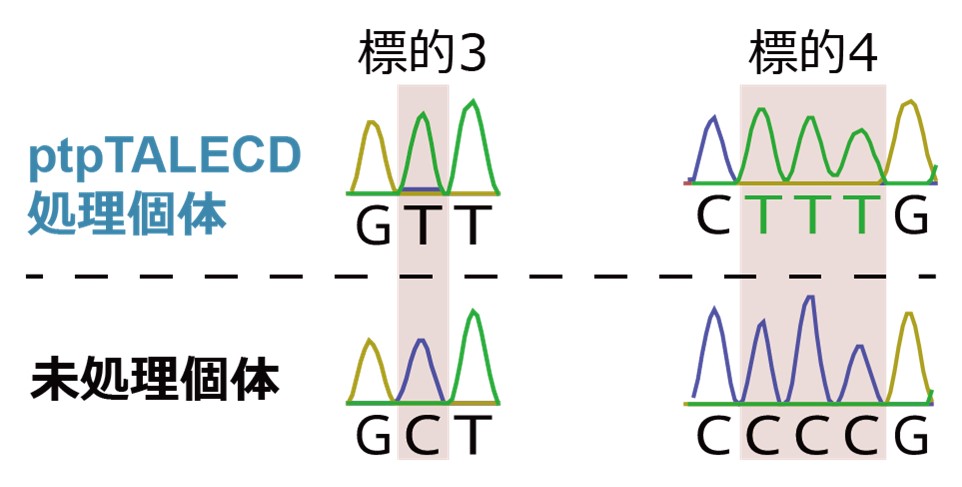
図3 ptpTALECDによる、GやCの直後のCの置換
ptpTALECD導入個体(上)では、難標的とされていたGやCの直後のCが細胞内の葉緑体ゲノムコピーの全てでTに置換された。
一般にゲノム編集技術を用いる際には、標的領域外に生じるハズレの変異 (オフターゲット変異) が問題になる場合があります。以前の研究で、ptpTALECDによって「ハズレの変異を持たず、かつ標的の塩基置換のみが置換された個体」を得られることが示されていました。本研究では、ptpTALECD_v2がハズレの変異を導入したかどうかを検証するため、植物体の葉緑体ゲノム全体の配列を解読しました。その結果、ptpTALECD_v2はptpTALECDと比べて酵素活性が高いためか、ハズレ変異の導入頻度も高くなっていることが分かりました。しかし、ハズレ変異が導入された個体の次世代の個体の中には、ハズレ変異が遺伝せず、しかし標的とした塩基が置換された葉緑体ゲノムが遺伝した個体が存在しました。これらの結果から、ptpTALECD_v2を用いて葉緑体ゲノムの標的塩基のみが置換された植物体を作出できることが示唆されました。
本研究では、ptpTALECD_v2はptpTALECDと比べて塩基置換活性が高い一方で、本当に置換したい塩基だけでなく標的以外の塩基も置換しやすいことが分かりました。そのため、標的塩基のみが置換された植物体を得るためには、まずptpTALECDによって標的塩基の置換を試み、それでも標的塩基を置換できなかった場合にptpTALECD_v2を用いるのが良いと考えられ、ゲノム編集酵素の選択肢を広げることができました。また、GやCの直後のCを置換できることが分かったため、葉緑体ゲノムのより多くのシトシンが置換の対象候補になりました。したがって、標的一塩基置換技術が葉緑体ゲノムの基礎研究に役立つことや、葉緑体ゲノムを利用した作物品種改良のための基盤技術になることが期待されます。
研究グループ
中里 一星(東京大学大学院農学生命科学研究科 生産・環境生物学専攻 博士課程2年/ 日本学術振興会特別研究員DC1)
奥野 未来(久留米大学医学部医学科 助教)
伊藤 武彦(東京工業大学 生命理工学院 教授)
堤 伸浩(東京大学大学院農学生命科学研究科 教授)
有村 慎一(東京大学大学院農学生命科学研究科 准教授)
発表雑誌
- 雑誌
- The Plant Journal
- 題名
- Characterization and development of a plastid genome base editor, ptpTALECD
- 著者
- Issei Nakazato, Miki Okuno, Takehiko Itoh, Nobuhiro Tsutsumi, Shin-ichi Arimura
- DOI
- 10.1111/tpj.16311
- URL
- https://onlinelibrary.wiley.com/doi/10.1111/tpj.16311
研究助成
本研究(の一部)は、日本学術振興会特別研究員奨励費22J20237,科研費基盤研究S 20H05680,国際共同研究加速基金(国際共同研究強化(A))19KK0391, JST -ASTEP JPMJTR22UGの支援を受けたものです。
用語解説
- 注1 モデル植物
生活環が短い、狭い空間で生育可能、ゲノムが解読済み、ゲノムが比較的単純で遺伝子操作しやすい等の理由により、研究でよく用いられる植物。シロイヌナズナは双子葉植物のモデル生物としてよく使用される。 - 注2 標的一塩基置換酵素ptpTALECD
ptpTALECDはplastid-targeted platinum TALE cytidine deaminaseの略。葉緑体ゲノム上の任意のDNA配列に結合し、その近傍のC:G対をT:A対に置換する酵素。特定のDNA配列に結合するTALEドメインや塩基置換を担うcytidine deaminaseドメインなどからなる。 - 注3 シトシン (C)、チミン (T)
核酸塩基の一種。DNAを構成する核酸塩基は主にアデニン (A)、チミン (T)、グアニン (G)、シトシン (C)の4種類。
問い合わせ先
〈研究に関する問合せ〉
東京大学大学院農学生命科学研究科 生産・環境生物学専攻
准教授 有村 慎一(ありむら しんいち)
E-mail: arimura[アット]g.ecc.u-tokyo.ac.jp
※[アット]を@に変えてください。