C4植物の光合成能力と環境適応力はC3植物よりも進化的に優れている――地球温暖化・気候変動に適応する植物の開発へ期待――
発表のポイント
- C4植物とC3植物において光の強さの変化に対する応答を調べたところ、C4植物、C3植物とC4植物の中間型、C3植物の順に光合成誘導と気孔応答が素早く、環境に素早く適応する能力を持っていることが分かりました。
- C4植物はC3植物から進化する過程でCO2濃縮機構という機能を獲得し、高温・乾燥地域に適応していると考えられています。この性質が、野外で日常的にさらされている変動光に対してどのように応答しているかを明らかにしました。
- 本研究成果は、植物が進化の過程で獲得した形質の有用性を証明するとともに、この優れた形質をほかの植物に導入し、将来の地球温暖化・気候変動に適応した植物を作出することの可能性を示しました。
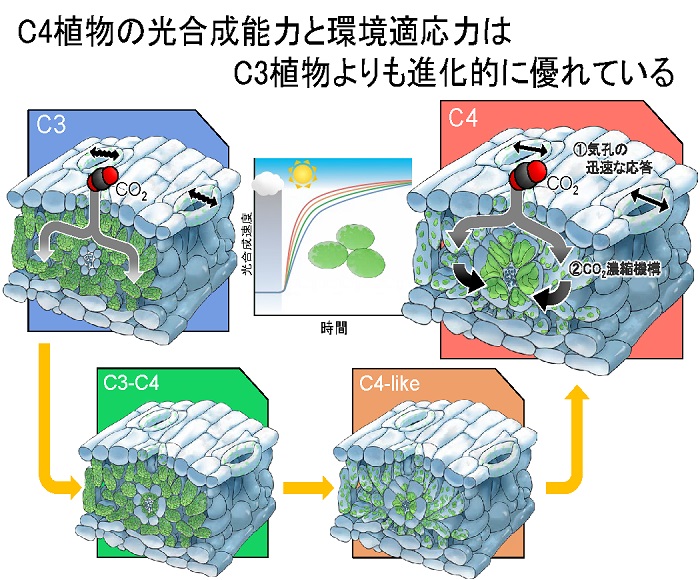
発表概要
東京大学大学院農学生命科学研究科の谷川慶一郎大学院生と矢守航准教授らは、C3植物、C4植物、それらの中間型を含むフラベリア属植物8種、イネ科植物2種の光合成特性を比較調査しました。その結果、弱光から強光に変化した際、C4植物、中間型、C3植物の順に素早く光合成が立ち上がることが分かりました。光合成に使用される二酸化炭素(CO2)は気孔(注1)を介して取り込まれますが、その気孔の開口もC4植物、中間型、C3植物の順に素早いことが明らかになりました。この結果から、C4植物は、CO2濃縮機構と素早い気孔応答という2つを活用して環境に適応するように進化したということが分かりました。
人類が食料として栽培している植物には、C3植物(イネやコムギなど)とC4植物(トウモロコシやサトウキビなど)の両方が含まれます。今後は、C4植物の優れた形質をC3植物に導入することによって、収量の向上や、地球温暖化や気候変動に対応できる耐環境ストレス品種の開発が期待されます。
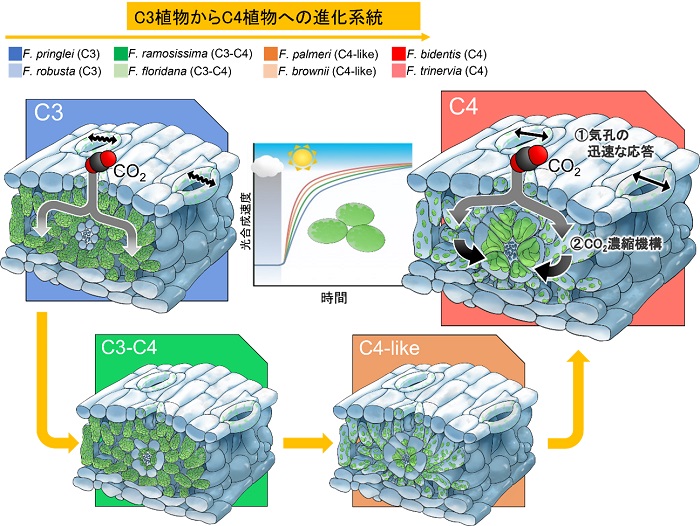
図1:C3植物からC4植物への進化系統
C4植物はC3植物からC3-C4中間型植物、C4-like植物を経て進化したと考えられており、その過程でCO2濃縮機構を獲得して高温・乾燥地域に適応しているとされています。我々は、これらの各段階の植物を比較調査することで、C4植物が迅速な気孔応答と、CO2濃縮機構を活かすことで素早く光に対して応答することができることを明らかにしました。
発表内容
昨今の地球温暖化とそれに伴う気候変動は作物の生育不良を引き起こしています。一方、世界の人口は増加し続けており、食料増産の必要性が叫ばれています。光合成は、植物が光からエネルギーを生産する営みであり、作物の収量を大きく左右する機能です。特に、C3植物から進化する過程でCO2濃縮機構という機能を獲得したことで高温・乾燥地域に適応しているC4植物は、C3植物には無い機能を持っている可能性があるため、C4植物の光合成の特徴を理解することは非常に重要です。
そこで東京大学大学院農学生命科学研究科の谷川慶一郎大学院生と矢守航准教授らは、C3植物、C4植物、それらの中間型を含むフラベリア属植物8種と、イネ科植物2種を栽培し、ガス交換測定装置LI-6400XTを用いて、それぞれの光合成特性を測定しました。CO2濃度を徐々に変化させながら光合成速度を測定した結果、CO2濃度が800 μmol mol-1以下では、フラベリア属・イネ科植物ともにC4植物がC3植物よりも優れた光合成速度を発揮することが分かりました(図2)。
次に、現在の大気CO2濃度に近いCO2濃度400 μmol mol-1および、CO2濃度が上昇した大気を模倣したCO2濃度800 μmol mol-1の2種類の環境下において、暗黒状態から一定の強光を照射した光合成誘導(注2)の解析を行いました。そして、光合成速度、気孔コンダクタンス(注3)、電子伝達速度(注4)が最大値の50%値に到達するまでに要した時間(= t50)と最大値の90%値に到達するまでに要した時間(= t90)を導出し、比較を行いました。結果として、CO2濃度400 μmol mol-1では、フラベリア属においてはC4植物、C4-like植物、C3-C4中間型植物、C3植物の順で、イネ科植物においてもC4植物、C3植物の順で、光合成速度と気孔コンダクタンスの素早い上昇がみられました(図3)。CO2濃度800 μmol mol-1では、C4植物の優位性は見られませんでした(図4)。
また、得られたデータから、CO2濃度400 μmol mol-1において光合成速度のt90とCO2補償点(= CO2濃縮機構の指標)、気孔コンダクタンスのt90とCO2補償点、光合成速度のt90と気孔コンダクタンスのt90の間には、それぞれ相関関係があることが判明しました(図5)。さらに、C4植物はCO2濃度に依存せず、常にC3植物より優れた水利用効率を示すことも明らかになりました(図6)。これらのことから、C4植物の素早い光合成誘導や乾燥に強いという特性は、進化の過程で獲得したCO2濃縮機構や素早い気孔応答に起因することが示唆されました。
本研究は、C4植物の光に対する優れた応答性を明らかにし、進化的な優位性を示した点で画期的です。また、本研究で調査した高濃度CO2環境下でのC3植物、C4植物の挙動は、これから人類が直面する地球温暖化、気候変動などの問題を解決するための重要な研究成果となるでしょう。
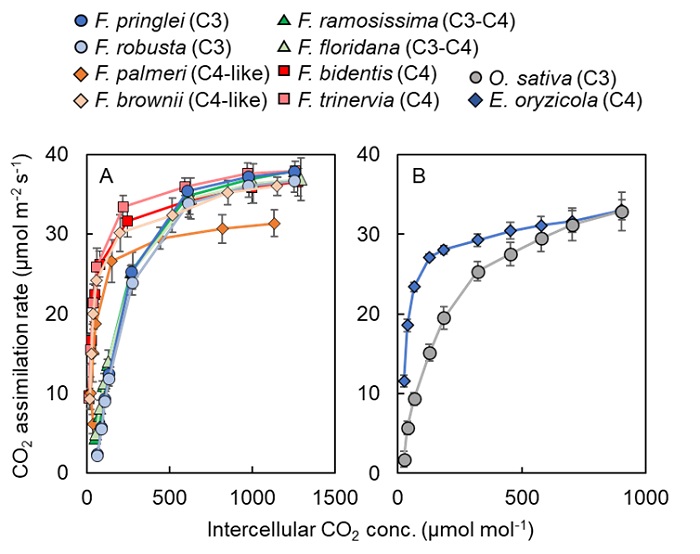
図2:光合成のCO2応答曲線
(A)フラベリア属8種における光合成のCO2応答曲線
(B)イネ科2種における光合成のCO2応答曲線
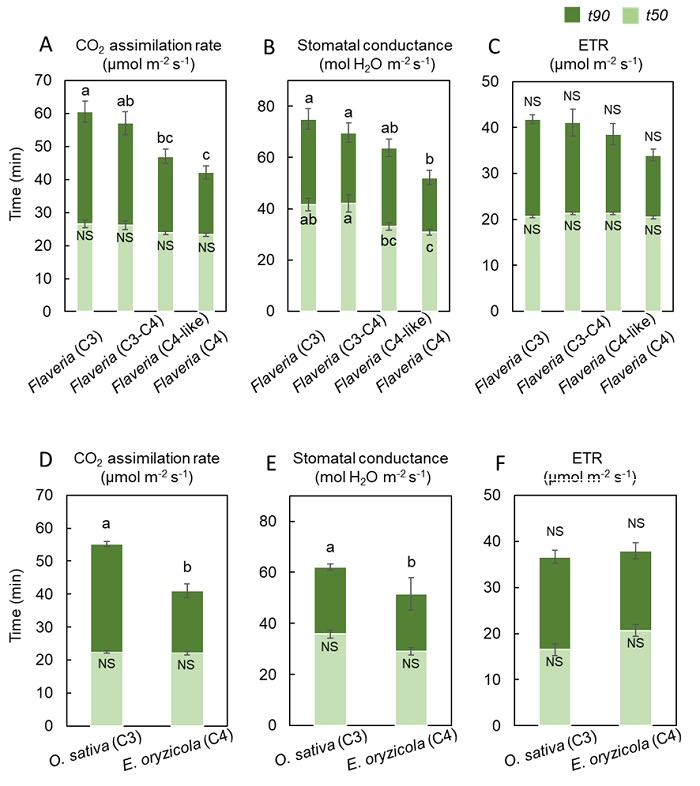
図3. CO2濃度400 μmol mol-1におけるフラベリア属とイネ科の光合成特性
CO2濃度400 μmol mol-1における (A)フラベリア属の光合成速度 (= CO2 assimilation rate)、(B)フラベリア属の気孔コンダクタンス (= Stomatal conductance)、(C)フラベリア属の電子伝達速度 (= ETR)、(D)イネ科の光合成速度、(E)イネ科の気孔コンダクタンス、(F)イネ科の電子伝達速度が、それぞれ最大値の50%値に到達するまでに要した時間 (= t50、薄緑色)と光合成速度の最大値の90%値に到達するまでに要した時間 (= t90, 緑色)。添付したアルファベット (a、b)が異なる種間では有意差があることを示す (P < 0.05)。
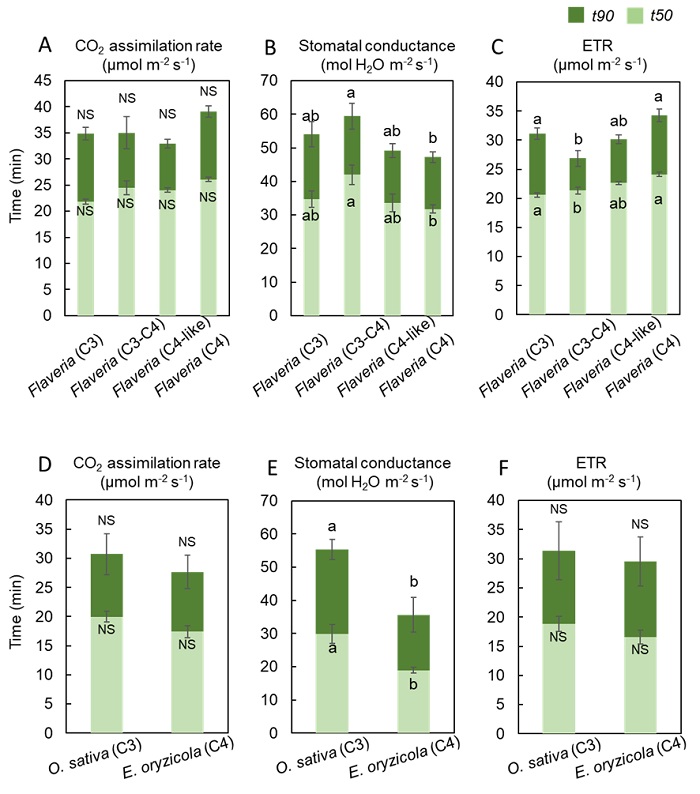
図4. CO2濃度800 μmol mol-1におけるフラベリア属とイネ科の光合成特性
CO2濃度400 μmol mol-1における (A)フラベリア属の光合成速度 (= CO2 assimilation rate)、(B)フラベリア属の気孔コンダクタンス (= Stomatal conductance)、(C)フラベリア属の電子伝達速度 (= ETR)、(D)イネ科の光合成速度、(E)イネ科の気孔コンダクタンス、(F)イネ科の電子伝達速度が、それぞれ最大値の50%値に到達するまでに要した時間 (= t50、薄緑色)と光合成速度の最大値の90%値に到達するまでに要した時間 (= t90, 緑色)。添付したアルファベット (a、b)が異なる種間では有意差があることを示す (P < 0.05)。
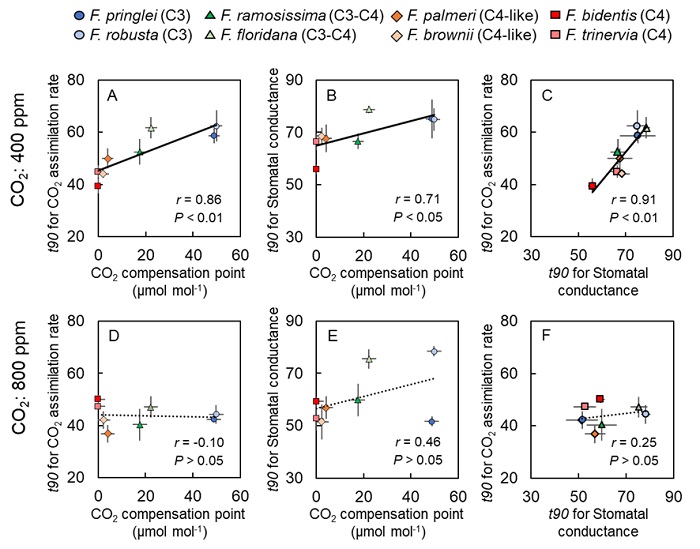
図5. 光合成誘導反応におけるフラベリア属の各パラメータの相関関係図
CO2濃度400 μmol mol-1におけるフラベリア属の (A)光合成速度のt90 (= t90 for CO2 assimilation rate)とCO2補償点 (= CO2濃縮機構の指標, CO2 compensation point)の相関関係、(B)気孔コンダクタンスのt90 (= t90 for Stomatal conductance)とCO2補償点の相関関係、(C)光合成速度のt90と気孔コンダクタンスのt90の相関関係、およびCO2濃度800 μmol mol-1におけるフラベリア属の (D)光合成速度のt90とCO2補償点の相関関係、(E)気孔コンダクタンスのt90とCO2補償点の相関関係、(F)光合成速度のt90と気孔コンダクタンスのt90の相関関係。近似直線はP < 0.05 の場合は実線で, P ≧ 0.05 の場合は点線で示されている。
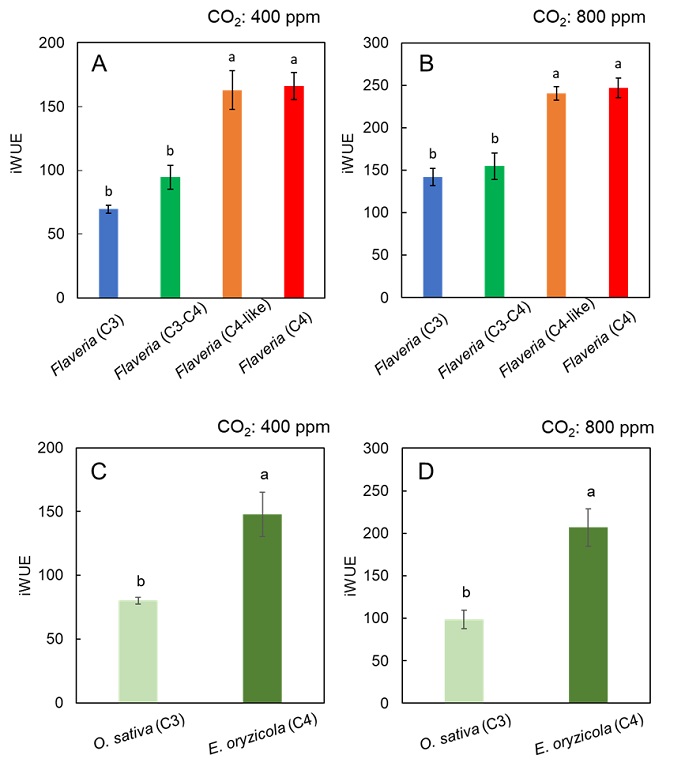
図6. フラベリア属とイネ科の水利用効率
フラベリア属の (A)CO2濃度400 μmol mol-1における水利用効率 (= iWUE)、(B)CO2濃度800 μmol mol-1における水利用効率、およびイネ科の (C)CO2濃度400 μmol mol-1における水利用効率、(D)CO2濃度800 μmol mol-1における水利用効率。添付したアルファベット (a, b)が異なる種間では有意差があることを示す (P < 0.05)。
発表者
東京大学大学院農学生命科学研究科
谷川 慶一郎 博士課程
Qu Yuchen 博士研究員
勝濵 直椰 博士課程
迫田 和馬 研究当時:博士研究員
若林 侑 助教
矢守 航 准教授
岡山大学環境生命自然科学学域
田中 佑 准教授
Department of Ecology and Evolutionary Biology, University of Toronto
Rowan Sage 教授
School of Life Sciences, University of Essex
Tracy Lawson 教授
発表雑誌
- 雑誌
- Phygiologia Plantarum
- 題名
- C4 monocots and C4 dicots exhibit rapid photosynthetic induction response in contrast to C3 plants
- 著者
- Keiichiro Tanigawa, Qu Yuchen, Naoya Katsuhama, Sakoda Kazuma, Yu Wakabayashi, Yu Tanaka, Rowan Sage, Tracy Lawson, Wataru Yamori
- DOI
- 10.1111/ppl.14431
- URL
- https://onlinelibrary.wiley.com/doi/full/10.1111/ppl.14431
研究助成
本研究は、日本学術振興会科研費「基盤研究(S)(課題番号:20H05687)」、「基盤研究(B)(課題番号:21H02171)」、「国際共同研究加速基金(国際共同研究強化(B))(課題番号:18KK0170)」、「学術変革領域研究(A)「細胞質ゲノム制御」(24A301、24H02277)」の支援により実施されました。
用語解説
- 注1 気孔
葉の表皮に主に存在する開閉式の小さい孔。周囲に位置する孔辺細胞という一対の細胞が膨張収縮することによって開閉し、植物体と大気間のガス交換 (CO2吸収や蒸散) を可能にしている。一般に夜間は不要な蒸散を防ぐべく閉口し、光照射とともに開口していく性質がある。 - 注2 光合成誘導
弱い光から強い光への急な変化に対して、光合成の速度は緩やかに上昇しながら最大値に達するという応答を示す。この応答を光合成誘導と呼ぶ。 - 注3 気孔コンダクタンス
大気から葉内への気孔を介したCO2輸送の効率を表す指標。 - 注4 電子伝達速度
光合成反応を構成する機能の一つであるチラコイド反応において、光のエネルギーによってどのくらいの化学エネルギーが生まれたかを表す指標。この指標で、光のエネルギーをどのくらい効率的に利用することができているかを推定することができる。
問い合わせ先
(研究内容については発表者にお問合せください)
東京大学大学院農学生命科学研究科附属生態調和農学機構
准教授 矢守 航(やもり わたる)
E-mail: yamori[at]g.ecc.u-tokyo.ac.jp
東京大学大学院農学生命科学研究科・農学部
事務部 総務課総務チーム 総務・広報情報担当(広報情報担当)
TEL: 03-5841-8179, 5484 FAX:03-5841-5028
E-mail: koho.a[at]gs.mail.u-tokyo.ac.jp
※上記の[at]は@に置き換えてください。