葉緑体ゲノム編集で除草剤に強い植物を作出 ――除草剤耐性作物の効率的生産に繋がる成果――
発表のポイント
- ゲノム編集技術を用いて、葉緑体ゲノムにコードされるD1タンパク質上の2つの標的アミノ酸置換の集積に成功し、市販の除草剤メトリブジンに強いシロイヌナズナを作出しました。
- 2アミノ酸が置換された個体は、1アミノ酸のみを置換した個体よりも耐性が高く、またメトリブジンなしの通常条件下でも非置換個体と同等に生育する優れた形質を示しました。
- 本研究で効果が実証されたアミノ酸置換の水平導入により多様な作物種へメトリブジン耐性の付与が可能であり、作物の安定生産への応用が期待されます。

ゲノム編集技術を用いて除草剤メトリブジンに耐性を示すシロイヌナズナの作出に成功!
発表概要
東京大学大学院農学生命科学研究科の中里一星大学院生と有村慎一教授らの研究グループは、除草剤メトリブジン(注1)が結合する葉緑体ゲノム上のD1タンパク質(注2)内の離れた2つのアミノ酸の置換・集積により、モデル植物シロイヌナズナ(注3) へのメトリブジン耐性の付与に成功しました。置換を集積した個体は1アミノ酸のみを置換した個体よりも強い耐性を示し、かつメトリブジンを含まない通常条件下でも非置換個体と同等に生育しました。他の手法では、離れた2アミノ酸の置換を引き起こすDNA変異の導入・集積はほぼ不可能でしたが、発表者らの開発した葉緑体ゲノム編集技術(注4)の利用により複数変異の集積が達成されました。メトリブジンが結合する領域のアミノ酸配列は多くの作物種で共通しているため、多様な作物種においても同様のアミノ酸置換の導入によるメトリブジン耐性品種の開発が可能と考えられます。将来的な作物の安定生産への貢献が期待される成果といえます。

図1:土壌で栽培した個体へのメトリブジン処理
作物栽培時に使用推奨される濃度と量のメトリブジンを散布した植物体(右側パネル)とメトリブジンの代わりに水を散布した個体(左側パネル)の様子。メトリブジン散布8日後において(右下パネル)、非置換個体は枯死したが、2アミノ酸を置換した個体は全12個体が生き残った。写真は全て同じ縮尺(バーの長さは1 cm)。
発表内容
除草剤は雑草を防除することで作物の安定生産に寄与してきましたが、作物(品)種の中には除草剤の働きで枯死するものもあります。これまで作物への除草剤耐性の付与は主に核ゲノムの改変により達成されてきましたが、核ゲノムに起因する除草剤耐性形質は次の世代の全ての個体には伝わらない場合があり、また花粉を介して作物と近縁な野生植物に導入されるリスクがあります。一方、葉緑体ゲノムは多くの作物種で母性遺伝するため、母親(種子親)の葉緑体ゲノムに除草剤耐性をもたらすDNA変異を導入すれば、その除草剤耐性形質は次の世代に100%遺伝し、かつ花粉を介した変異拡散がありません。しかし、葉緑体ゲノムを利用した除草剤耐性作物の育種はこれまでほとんど行われてきませんでした。その要因として、葉緑体ゲノム改変技術(外来遺伝子導入法)が限られた植物種にしか適用できないことや、作出された植物体が遺伝子組換え生物として扱われることから野外栽培への大きな制約があることが挙げられます。
本研究では、葉緑体ゲノム編集酵素ptpTALECDとptpTALECD_v2(注6)を用いてD1タンパク質に2つのアミノ酸置換(219番目のバリンをイソロイシンに、251番目のアラニンをバリンに置換) を導入することで、メトリブジンに強い耐性を持つシロイヌナズナの開発に成功しました。この2アミノ酸が置換された個体は、1アミノ酸のみが置換された個体よりも強い耐性を示し、また作物栽培時の使用推奨濃度および量のメトリブジン散布によっても生存し(図1)、さらにメトリブジンを処理しない条件では非置換個体と同等の生育を示しました(図2)。D1タンパク質においてメトリブジンが結合する領域のアミノ酸配列は、219番目のバリンや251番目のバリンを含め、多くの作物種で共通していることから、これらアミノ酸置換の水平導入による他作物へのメトリブジン耐性の付与は、有効性と安全性が認められたメトリブジンの適用可能な作物種の拡大に繋がります。さらに、今回利用したゲノム編集技術は、最終的にゲノム中に外来遺伝子を含まない個体の作出が可能であり、かつゲノム編集技術以外の方法によるこれら2つのアミノ酸置換をもたらすDNA変異の導入は困難なことから、ptpTALECDやptpTALECD_v2を用いた葉緑体ゲノム編集技術はメトリブジン耐性作物育種のための有効な技術になることが期待されます。
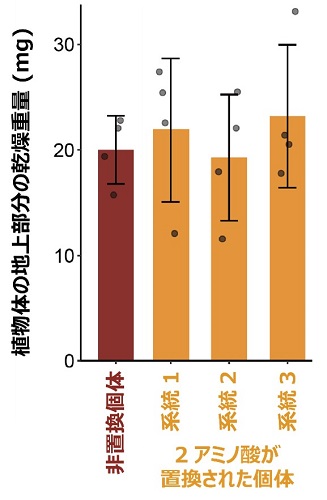
図2:土壌で栽培した未置換個体と2アミノ酸を置換した個体の地上部分の乾燥重量
植物の生育の指標の一つである地上部分の乾燥重量は、メトリブジンを処理しない場合、非置換個体と2アミノ酸を置換した個体で統計的に有意な差が無かった。その他の生育の指標(根の乾燥重量、サヤの乾燥重量、葉の面積など)についても統計的に有意な差が無かった。
発表者
東京大学 大学院農学生命科学研究科
有村 慎一 教授
堤 伸浩 教授
矢守 航 准教授
Qu Yuchen 特任研究員
中里 一星 大学院生 兼 日本学術振興会特別研究員
立命館大学 生命科学部
松村 浩由 教授
久留米大学 医学部
奥野 未来 講師
論文情報
- 雑誌
- Plant Biotechnology Journal
- 題名
- Resistance to the herbicide metribuzin conferred to Arabidopsis thaliana by targeted base editing of the chloroplast genome
- 著者
- Issei Nakazato, Wataru Yamori, Hiroyoshi Matsumura, Yuchen Qu, Miki Okuno, Nobuhiro Tsutsumi, Shin-ichi Arimura*
- DOI
- doi/10.1111/pbi.14490
- URL
- https://onlinelibrary.wiley.com/doi/10.1111/pbi.14490
用語解説
- 注1 メトリブジン
D1タンパク質に結合することで光合成反応を阻害し、植物を枯死させる除草剤。日本、北米、欧州、インド、オーストラリア等で市販・利用されている。 - 注2 D1タンパク質
光合成反応に必要な電子伝達を媒介するタンパク質。 - 注3 モデル植物シロイヌナズナ
「個体サイズが小さいため少ないスペースで栽培可能」、「ゲノム情報が解読されており、かつゲノム構造が複雑でない」、「核ゲノムに外来遺伝子を導入可能」、「世代時間が短い」といった理由から植物研究のモデル生物として世界中で用いられている植物。 - 注4 ゲノム編集技術
最終的にゲノム中に外来遺伝子断片を残すことなく、ゲノムの狙った配列を意図した通りに改変する技術。ゲノムとは、生物が保持する遺伝情報の総体のこと。 - 注5 母性遺伝
母親のゲノムのみが次の世代に伝わること。 - 注6 ptpTALECD、ptpTALECD_v2
発表者らが以前に開発したゲノム編集酵素で、葉緑体ゲノム上の狙ったC:G対をT:A対に置換する。A, T, G, CはDNA塩基の略号であり、それぞれアデニン、チミン、グアニン、シトシンを指す。
研究助成
本研究は、科研費「基盤研究S(課題番号:20H00417)」、「研究拠点形成事業(課題番号:JPJSCCA20230008)」、「特別研究員奨励費(課題番号:22J20237)」、「学術変革領域研究(A)「細胞質ゲノム制御」(課題番号:24H02271; 24H02277)」、JST「ASTEP(課題番号:JPMJTR22UG)」の支援により実施されました。
問い合わせ先
東京大学大学院農学生命科学研究科
教授 有村慎一 (ありむら しんいち)
E-mail:arimura[at]g.ecc.u-tokyo.ac.jp
※上記の[at]は@に置き換えてください。
関連教員
有村 慎一
堤 伸浩
矢守 航