日本のモウソウチクは、種子ではなく、 株か地下茎(栄養体)で、海を越えて伝播してきた! ――ヘテロ接合箇所に着目したゲノム解析による推定――
発表のポイント
- 中国15地域由来のモウソウチクと鹿児島に渡来したと伝わる日本のモウソウチクの全ゲノム配列解析により、解析した全サンプルはクローン(単一個体由来の栄養生殖個体)であることを明らかにしました。
- ヘテロ接合座位に着目した新規ゲノム解析手法により、文献記録や従来解析手法では不明であった系統関係を明らかにし、中国・福建省の竹が日本に由来したと推定しました。
- 本発表はクロ―ナル植物のゲノム多様性に関する重要な新規な知見であり、この新規解析手法は他のクロ―ナル植物のゲノム多様性研究への応用が期待されます。

発表内容
〈研究の背景〉
モウソウチクは東アジアに広く分布する竹です。日本のモウソウチクは江戸時代に中国から鹿児島にはじめて導入されたと考えられていますが、中国のどの地域からどのように持ち込まれたのかは不明です。
モウソウチクは、67年前後の期間をあけての開花例が知られるものの、通常花を咲かせず、地下茎の伸長や人の手による移植によって分布を拡げます。その為、ゲノム(注1)の多様性が小さいと考えられます。花を咲かせず、交配しなくても、稀にヘテロに生じた突然変異によりゲノムの多様性が生じ、その変異が拡散すると考えられますが、これまでの少数のDNAマーカー(注2)を用いた解析ではゲノムの多様性を十分に評価することができませんでした。
しかし、近年モウソウチクのゲノム配列が解読され、その概要が明らかになりました。そして、このゲノム情報を参照して、様々な系統のゲノムの比較解析を行うことができるようになりました。 そこで、東京大学 大学院 農学生命科学研究科の大学院生である西山典秀と教授の井澤毅の研究チームは、東京農業大学と共同研究し、ショートリード型次世代シークエンサー(注3)を用いたゲノム解析を行うことで、モウソウチクがどのようなゲノム多様性を持つのか、日本のモウソウチクの由来はどうなっているのか、モウソウチクは花を咲かせずに分布を拡げる中でどのようなゲノムの多様性を獲得するのか、等を明らかにすることを目指して研究を進めました。
〈研究の内容〉
まず、江戸時代に日本に最初に導入されたと考えられているモウソウチク1系統と、モウソウチクの稈に奇形を生じた品種であるキッコウチクの日本の2系統を、新規に全ゲノム配列解読しました。次に、それらと中国各地から均等に選ばれた15系統のモウソウチクの既報の全ゲノム配列データを用いて、全ゲノム配列の比較解析を行いました。今回、これまでは信頼度が低いと考えられ、あまり解析に使われてこなかったヘテロ接合箇所と判断された座位を、評価基準を厳しく設定することで注目し、解析を行いました。
有性生殖により種子を経ると、親の世代のヘテロ接合(注4)座位の一部はメンデルの法則に従って後代でホモ接合になりますが、解析したサンプルと参照ゲノムとの間で検出されたSNP(注5)の大多数(77%)は、全サンプル共通のヘテロ接合でした(図1)。このことから、今回、解析された全サンプルは単一クローン由来の栄養繁殖個体と推定しました。したがって、解析した中国全体に分布するモウソウチクはクローンと考えられ、日本に渡来したモウソウチクも、種子ではなく、地下茎等の栄養体で海を越えて伝わってきたと考えられます。
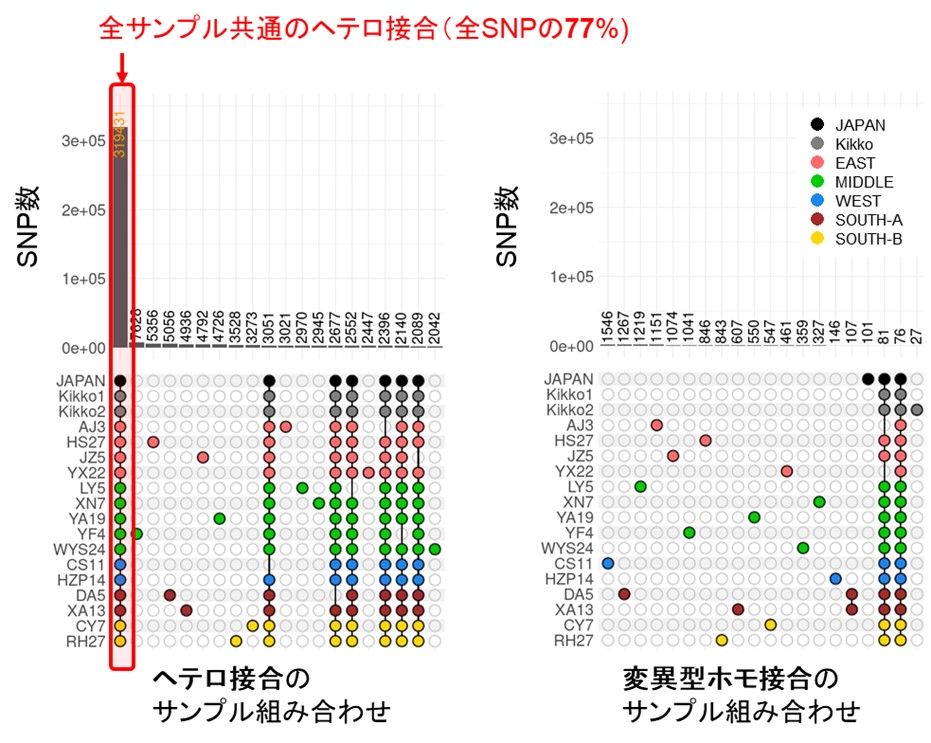
図1 中国、日本の全18サンプルのモウソウチクのヘテロ接合、変異型ホモ接合のサンプル組み合わせ
縦軸のJAPAN~RH27はサンプル名を表す。JAPANは日本に最初に導入されたと考えられているモウソウチクの系統。Kikkoは日本のキッコウチク、EAST、MIDDLE、WEST、SOUTH-A、SOUTH-Bは中国サンプルの地域分類。 「ヘテロ接合のサンプル組み合わせ」、「変異型ホモ接合のサンプル組み合わせ」ともに、上位20の組み合わせのみを図示。
更にサンプル間で共通するヘテロ接合の座位に注目した解析を行うことで、従来のゲノム解析手法では不明であった系統関係の推定が可能となりました。この解析により、福建省のサンプルに近縁の栄養生殖個体が海を越えて日本に導入されたと推定しました(図2、3)。
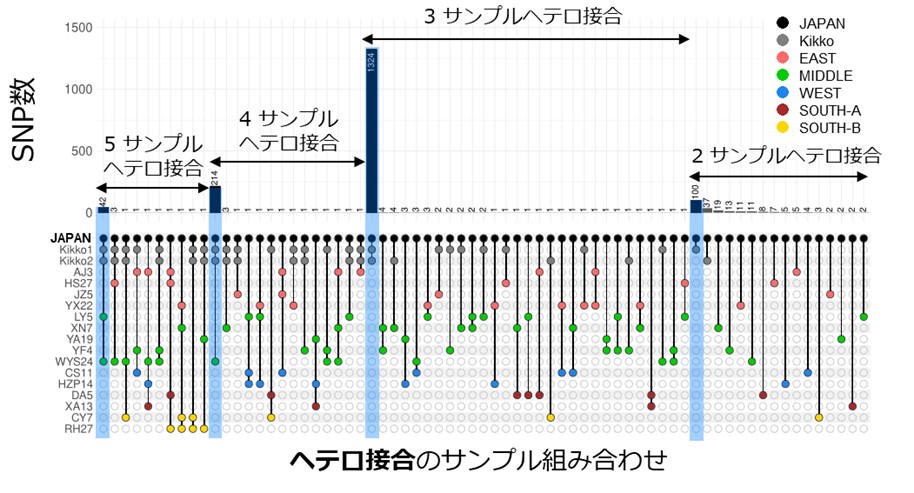
図2 ヘテロ接合に着目した解析による、日本のモウソウチクの由来の推定
飛びぬけたピーク(背景水色)が系統関係を示す。2~5サンプルがヘテロ接合の場合のみ図示。WYS24が福建省のサンプルであり、中国のサンプルの中で日本のモウソウチクと最も多く共通のヘテロ接合を持つ。
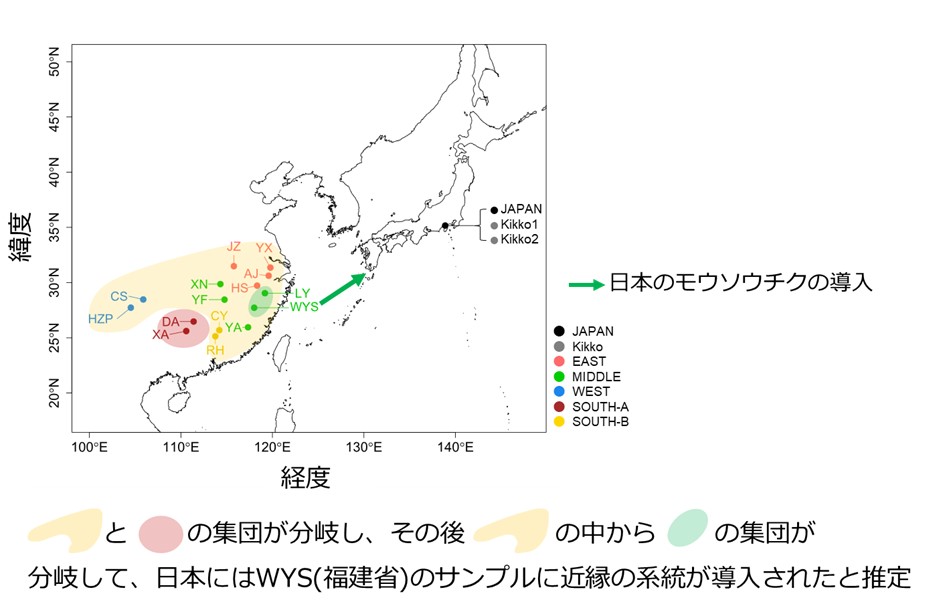
図3 推定された日本のモウソウチクの由来
また、ゲノム上のヘテロ接合の分布の偏りがあることから、現在中国から日本に広く分布するモウソウチクは、近縁のマダケ属間での交配により生じたF1個体由来ではなく、F2かF3世代の個体が広がったと考えられます。また、日本での極稀な開花の際に得られた種から育てた後代のモウソウチクでは、偏ったヘテロ接合の座位がメンデルの法則に従う形でホモ接合の座位に置き換わっていることから、解析結果の確からしさが裏付けられました(図4)。
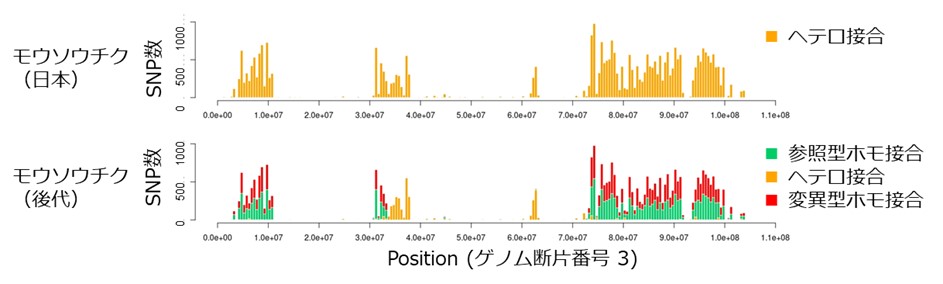
図4 現在中国から日本に広く分布するモウソウチクの系統は、過去に交配を経験したと推定
モウソウチクのヘテロ接合座位は局在していた(図上)。種子由来のモウソウチク(図下)では、図上でヘテロ接合の多くがホモ接合に置き換わっている。一本のバーは,1Mbpごとの集計を示すが、参照型と変異型が混在しているのは、参照したゲノム配列が1つの個体が持つ2つのゲノム配列セット(ハプロタイプ)を考慮せずに作成されていることを反映していると考えられる。
次に、静岡県内の4つの近接する竹林から、それぞれ4サンプルずつ異なる稈からサンプルを採取し、GRAS-Di(注6)を用いてサンプル間のSNPInDel(注7)を調査しました。栄養生殖の過程で生じるゲノム多様性は、突然変異によるヘテロ接合と考えられるため、参照ゲノム型ホモ接合とヘテロ接合のサンプルの両方のみを含むlociのみを抽出し、ゲノムビューアー(注8)による人による目視検査や、サンガーシークエンス(注9)を行うことで、真のDNA変異を特定しました。その結果、6.1Mbp(全ゲノム1.9 Gbの約 0.32%の解析領域)において、7 lociのヘテロ接合のSNP、2 lociの変異型ホモ接合のSNPInDelのみが確認されました(図5)。これらのDNA変異は、中国の15地域のサンプルでは見つからなかった為、モウソウチクが日本に導入されてから生じた可能性が考えられました。また、同一竹林内や近隣の竹林で、変異が定着したと考えられるDNA変異も見つかったことから、突然変異により生じた少数のヘテロ接合が、竹林内で定着し、地下茎の伸長や人の手による移植等によって広がったと考えられます。また、予想に反して見つかった変異型ホモ接合は、突然変異で生じたヘテロ接合の遺伝子座において、2本鎖切断と相同組換え修復(注10)のある種のDNA修復で生じたと推測しました。
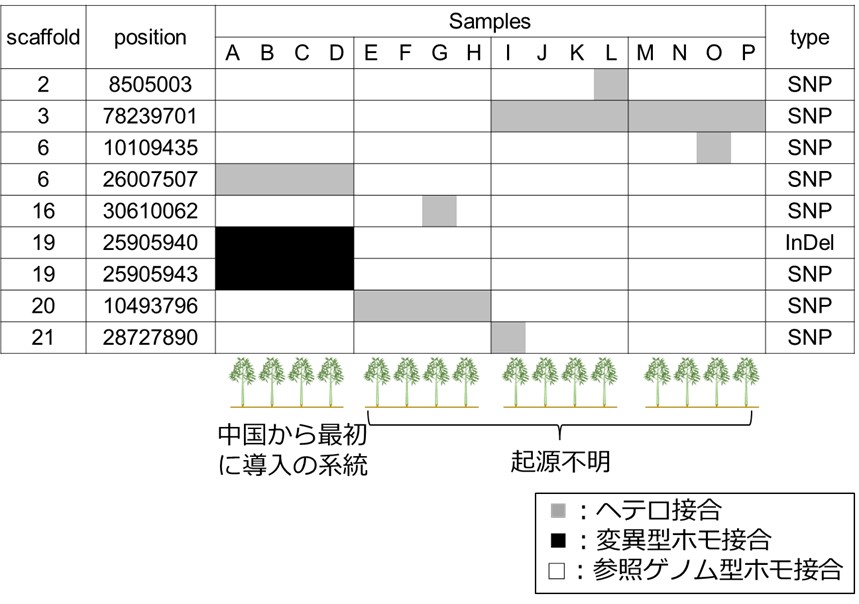
図5 日本の4つの竹林内で見つかった9つのサンプル間SNPInDel
加えて、GRAS-Diデータを用いた今回の解析においては、PCR(注11)増幅時のバイアスや、参照ゲノムの不完全性、ゲノムのヘテロ接合性により誤った遺伝子型判定のリスクがあり、ゲノムビューアーを用いた目視検査やサンガーシークエンスでの確認を行うことが重要であることが明らかになりました。
〈今後の展望〉
本発表のモウソウチクのゲノムの多様性に関する新たな知見は、クロ―ナル植物の生存戦略や、有性生殖の意義に関する重要な発見です。今後、静岡以外の日本中に分布するモウソウチクがクローンか種子を経て増えているかどうかを確認していく予定です。またゲノム配列のヘテロ接合に着目した本研究で記載した新規の系統解析手法は、他のクロ―ナル植物のゲノム多様性研究への応用が期待されます。
〈関連のプレスリリース〉
「日本育種学会 第142回講演会記者発表」(2022/09/14)
https://jsbreeding.jp/2022/09/15/post-677/
研究グループ
東京大学大学院農学生命科学研究科
西山 典秀(博士課程)
井澤 毅(教授)
東京農業大学・バイオサイエンス学科
篠澤 章久(助教)
松本 隆(教授:当時)
発表雑誌
- 雑誌
- BMC Genomics
- 題名
- High genome heterozygosity revealed vegetative propagation over the sea in Moso bamboo
- 著者
- Norihide Nishiyama, Akihisa Shinozawa, Takashi Matsumoto, Takeshi Izawa*(*責任著者)
- URL
- https://bmcgenomics.biomedcentral.com/articles/10.1186/s12864-023-09428-9
研究助成
本研究(の一部)は、Human Frontier Science Program Organization (grant number RGP0011/2019)、内閣府ムーンショット型農林水産研究開発事業(管理法人:生研支援センター)「JPJ009237」、東京農業大学生物資源ゲノム解析センターの生物資源ゲノム解析拠点事業における共同利用・共同研究、科研費 学術変革A「挑戦的両性花原理」JP22H05180&JP22H05172 、JST 次世代研究者挑戦的研究プログラム JPMJSP2108、の支援を受けたものです。
用語解説
- 注1 ゲノム
DNA の文字情報に表された遺伝情報の全て。A、T、G、C の 4 種類の塩基と呼ばれる部品から構成される。モウソウチクのゲノムは約 19 億塩基対で構成されており、シロイヌナズナ(約1 億3 千万塩基対)、イネ(約 3 億 8 千万対)といった実験植物と比べてサイズが大きく、解析の難度とコストが共に高い。 - 注2 DNAマーカー
DNAの塩基配列上の特定の位置の個体間の差を判別する目印 - 注3 ショートリード型次世代シークエンサー
ゲノムDNAの塩基配列を、全長の数十倍に相当する大量の150bp長のDNA配列データとしてゲノムを解読する手法 得られた150bp長DNA配列を参照ゲノム配列にマップして、比較解析を行う。 - 注4 ヘテロ接合
多くの生物は、両親から 1 セットずつのゲノム情報を受け取る。この両親からの配列が異なる配列になっている座位を含む相同領域をヘテロ接合の状態と呼ぶ。自家受粉で種子ができて繁殖するケースでは、半分がヘテロな状態で、残りの 1/4 ずつが、片親と同じ配列(ホモ接合)になる。他殖の場合は、より複雑な割合で、いろいろな組み合わせができる。どちらを考慮しても、ヘテロ状態の領域は、交配により種子を経て、後代を作ると、後代では、ホモな状態の領域が増えていくことになる。 - 注5 SNP
single nucleotide polymorphism の略。1 塩基多型。突然変異等により生じた塩基配列の違いで、系統や種を判別するのに用いられる。 - 注6 GRAS-Di
GRAS-Di(Genotyping by Random Amplicon Sequencing-Direct)。ランダムプライマーを用いたPCR増幅により、ゲノムの複数箇所を配列決定し、遺伝子型を決定する手法。 - 注7 DNAマーカー
DNAの塩基配列上の特定の位置の個体間の差を判別する目印 - 注8 InDel
塩基の挿入(insertion)と塩基の欠失(deletion)を指す。SNPと同様に突然変異等により生じる塩基配列の違いで、系統や種を判別するのに用いられる。 - 注9 サンガーシークエンス
1970年代にフレデリック・サンガーが開発した塩基配列の決定法。ショートリードシークエンサーによる塩基配列決定よりも大幅にコストは高いが、信頼度の高い結果が得られる。 - 注10 相同組換え修復
何かしらの原因で切断されたDNAを、もう一対の正常な DNA を鋳型にしてコピーし、最後に組換えによって元に戻す修復機構。 - 注11 PCR
ポリメラーゼ連鎖反応(Polymerase Chain Reaction)。Primerと呼ばれる短いDNAや酵素を用いて、目的のDNA領域を複製して、増幅させる方法。
問い合わせ先
〈研究に関する問合せ〉
東京大学大学院農学生命科学研究科 生産・環境生物学専攻
教授 井澤 毅(いざわ たけし)
Tel:03-5841-5063 E-mail:takeshizawa[アット]g.ecc.u-tokyo.ac.jp
〈報道に関する問合せ〉
東京大学大学院農学生命科学研究科・農学部
事務部 総務課総務チーム 総務・広報情報担当(広報情報担当)
Tel: 03-5841-8179 Fax:03-5841-5028
E-mail:koho.a[アット]gs.mail.u-tokyo.ac.jp
※[アット]を@に変えてください。