ゼニゴケは自身の成長制御に植物ホルモン『ジベレリン』を使わず・作らず、ヒメツリガネゴケの制御物質とも構造が異なる『もっとジベレリンに似た分子』を使う
発表のポイント
- ゼニゴケは自身の形を制御するため、植物ホルモン・ジベレリンの原料から「ジベレリンとは別の活性物質(GAMP)」を作り、利用している。
- 過去、ヒメツリガネゴケから「ジベレリンとは別の活性物質」を見つけているが、ゼニゴケのGAMPはジベレリンに近い別の構造をしている。
- ゼニゴケのこの系にジベレリンを与えても、形が変化しない。
発表概要
草丈や発芽をコントロールすることで知られる植物ホルモン・ジベレリンは、数段階の過程を経て植物体内で合成されている【図1】。コケ植物には、蘚類や苔類という小分類【図2】があり、ゼニゴケは苔類、ヒメツリガネゴケは蘚類に属する。以前の研究[1]から、ヒメツリガネゴケはent-カウレン酸(KA)まで合成するが、ジベレリン合成に必要な以降の酵素遺伝子群が見つからず、体内からもジベレリンを検出できない。代わりに、KAの代謝物3OH-KAが分化制御に関わる生理活性物質と判明している。今回の研究により、ゼニゴケではヒメツリガネゴケで見つからなかった酵素遺伝子群の一部が存在し機能するため、KAからジベレリン前駆体GA12まで合成されていた。よって、ヒメツリガネゴケと同様にゼニゴケもジベレリンを合成しないものの、前駆体GA12もしくはその代謝物までは合成し、制御物質として使っていることを発見した。進化的にコケ植物よりも後に出現したすべての維管束植物(シダ植物や顕花植物)ではジベレリンを合成して使っており、ゼニゴケ・ヒメツリガネゴケいずれのコケ植物ともにジベレリンを使わずに別構造の物質を生命現象の制御に利用しているとは、何が契機となり一体ジベレリンはどこからきたか謎であり興味深い。ヒメツリガネゴケ3OH-KA、ゼニゴケGAMPは、ジベレリンの起源となる物質やその制御機構に思いを馳せる上で、大変重要なヒントを提供しているはずで、研究の進展に注目されたい。
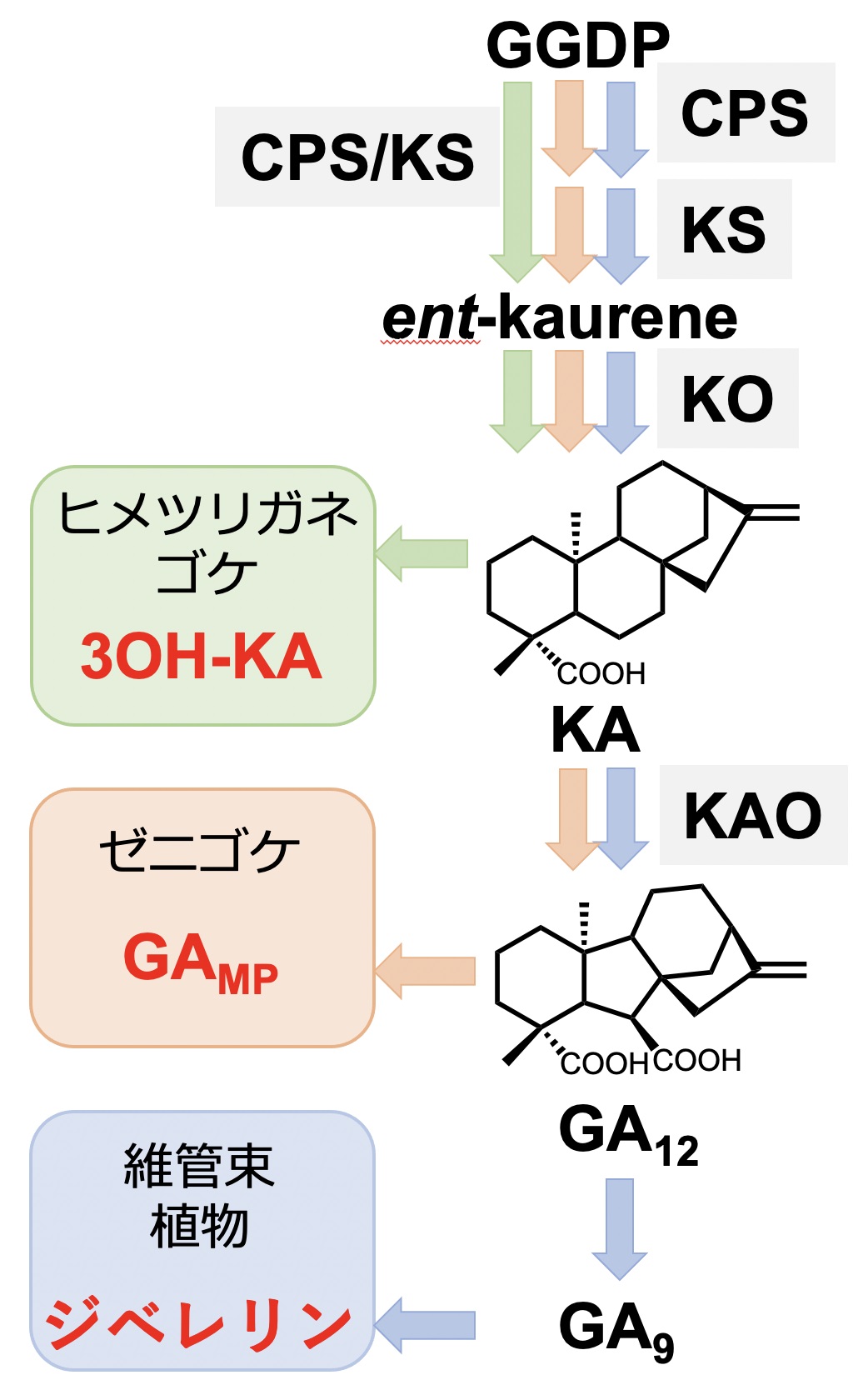
【図1】維管束植物(シダ植物や顕花植物)では、ゲラニルゲラニル2リン酸(GGPP)を原料として、説明文中に出てくる各種酵素群(CPS、 KS、 KO、 KAOなど)の触媒反応を経て「ジベレリン」は生合成されている。その経路途上には、本研究において重要な意味を持つKAやGA12など生合成中間体が位置する。先行研究のヒメツリガネゴケでは、KAは体内から検出されるが、KAO以降の酵素の存在が確認できずジベレリンも検出されない。その代わり、KAから3OH-KAへの代謝が確認され、分化制御に使われている。今回の研究により、ゼニゴケではKAO酵素が存在してGA12も検出されたが、以降の必要酵素は依然確認できずジベレリンも検出されない。GA12からの代謝物がGAMPの主要部分を担うと考えられる。
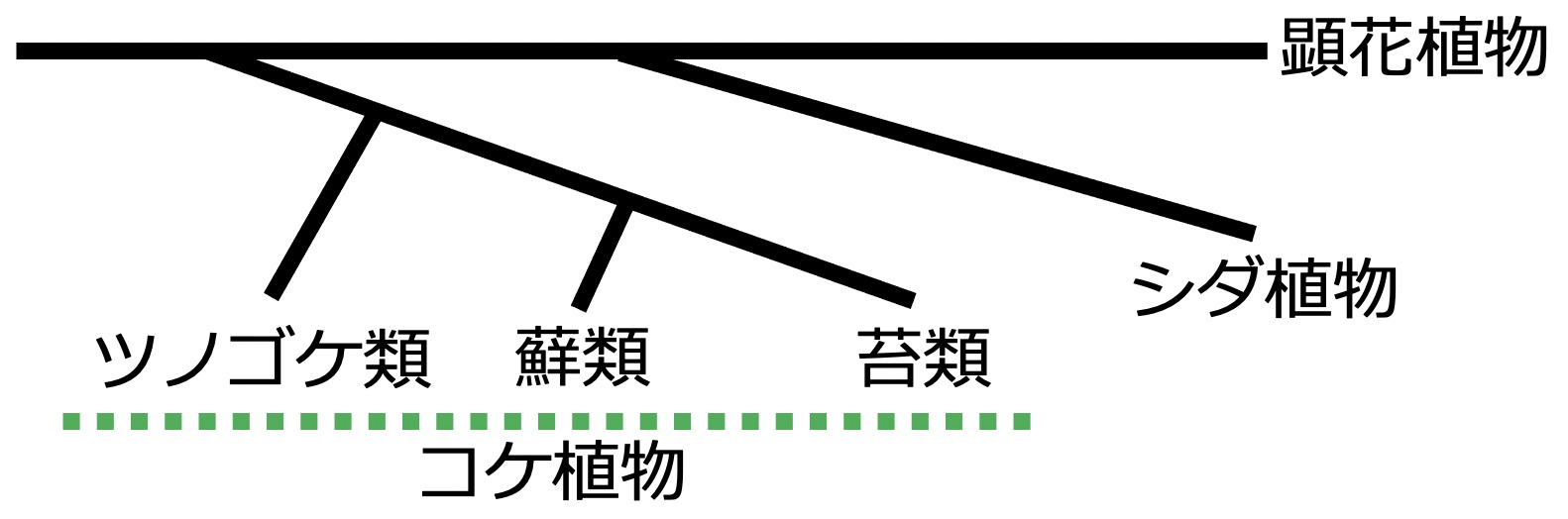
【図2】植物の進化過程において、コケ植物はツノゴケ類、蘚類、苔類の3つに大別されている。このうち、ゼニゴケは苔類に属し、ヒメツリガネゴケは蘚類に属する。
発表内容
ゼニゴケの生育に対するジベレリンの関与を判断すべく、カウレン合成能欠損変異体(cps欠損変異体)を作成したところ、頂端分裂組織数の増加、葉状体面積の拡大、遠赤色光で誘導される下偏成長の異常を認めた。このcps欠損変異体に対してCPS遺伝子の相補的発現により異常からの回復傾向を認め、これらの異常がCPSの機能欠損に起因することを把握した。 次に、LC-MS/MSを用いてジベレリンおよびその生合成前駆体のゼニゴケ内生量を分析した。結果、経路上の前駆体GA12のみ野生株から検出された。一方、cps欠損変異体ではGA12すら検出されなかった。cps欠損変異体では生合成過程上流の遮断に伴い、GA12だけでなくその原料KAなどが合成できない状態と考えられた。そこで、cps欠損変異体に対する外部投与実験を行ったところ、KA投与で異常からの回復傾向を認めたのに対し、ジベレリン投与では回復傾向を認めなかった。この結果より、ゼニゴケにおいてもヒメツリガネゴケと同様に、遠赤色光で誘導される下偏成長の制御に関わる活性代謝物(GAMPと呼称)がKAあるいはGA12から生合成され、使われていると解釈できる。 ゼニゴケにおけるGA12生合成に必要となる酵素遺伝子群(カウレン酸化酵素(KO)およびカウレン酸酸化酵素(KAO))は、いずれも触媒機能を持つ酵素をコードしているか検証すべく、KOとKAO両候補遺伝子を特定して酵母を用いた各組換え酵素を調製した。試験管内で各基質を用いた酵素反応を行ったところ、期待どおりカウレンからカウレン酸(KA)、KAからGA12への各変換活性を検出した。特定したKOおよびKAOの各酵素遺伝子に加え、さらに上流のカウレン合成酵素(KS)遺伝子を対象とした欠損変異ゼニゴケを作出したところ、前出のcps欠損変異体と同様に、遠赤色光で誘導される下偏成長に異常を確認した。ks欠損変異体およびko欠損変異体で認められた異常は、KA外部投与で回復した。しかし、kao欠損変異体の異常はKA外部投与で若干の変化のみを認めた。このことから、活性物質GAMPとしてKA自身か、GA12とは異なるKA代謝物によっても部分的ながら生理機能を果たし得ることを確認するとともに、GA12か、GA9と異なるGA12代謝物が主たるGAMPと考えている。
発表者
宮崎 翔(東京大学 大学院農学生命科学研究科 応用生命化学専攻 特任助教・当時、現・東京農工大学 グローバルイノベーション研究院 助教)
中嶋 正敏(東京大学 大学院農学生命科学研究科 応用生命化学専攻 准教授)
発表雑誌
- 雑誌
- The Plant Cell
- 題名
- Biosynthesis of gibberellin-related compounds modulates far-red light responses in the liverwort Marchantia polymorpha
- 著者
- R. Sun、 M. Okabe、 S. Miyazaki、 T. Ishida、 K. Mashiguchi、 K. Inoue、 Y. Yoshitake、 S. Yamaoka、 R. Nishihama、 H. Kawaide、 M. Nakajima、 S. Yamaguchi、 and T. Kohchi*
- DOI
- doi.org/10.1093/plcell/koad216
研究助成
本研究は、日本学術振興会科学研究費補助金 (課題番号:21H04722、22K19168、22KJ1083、23K13864) の支援を受けて行われました。
問い合わせ先
〈研究に関する問合せ〉
東京大学 大学院農学生命科学研究科 応用生命化学専攻
准教授 中嶋 正敏 (なかじま まさとし)
Tel:03-5841-5192 E-mail: anakajm[アット]g.ecc.u-tokyo.ac.jp
〈報道に関する問合せ〉
東京大学大学院農学生命科学研究科・農学部
事務部 総務課総務チーム 総務・広報情報担当(広報情報担当)
Tel: 03-5841-8179 Fax: 03-5841-5028
E-mail: koho.a[アット]gs.mail.u-tokyo.ac.jp
※[アット]を@に変えてください。
参考文献
[1] S. Miyazaki、 M. Hara、 S. Ito、 K. Tanaka、 T. Asami、 K. Hayashi、 H. Kawaide and M. Nakajima*. An ancestral gibberellin biosynthetic pathway in the moss Physcomitrella patens. Mol. Plant、 11: 1097-1100、 (2018). [doi: 10.1016/j.molp.2018.03.010].