栄養屈性により曲がる根の内側と外側で見られる植物ホルモン応答
- 発表者
- 山崎 清志(東京大学 大学院農学生命科学研究科 応用生命科学専攻: 特任講師)
大森 良弘(東京大学 大学院農学生命科学研究科 アグリバイオインフォマティクス教育研究ユニット: 准教授)
高橋 宏和(名古屋大学 大学院生命農学研究科 植物生産科学専攻: 准教授)
豊田 敦(国立遺伝学研究所 先端ゲノミクス推進センター: 特任教授)
佐藤 豊(国立遺伝学研究所 ゲノム・進化研究系: 教授)
中園 幹生(名古屋大学 大学院生命農学研究科 植物生産科学専攻: 教授)
藤原 徹(東京大学 大学院農学生命科学研究科 応用生命科学専攻: 教授)
発表のポイント
植物の根が栄養の濃度勾配に曝されると、栄養の濃い方向に根の伸長方向を変えます(栄養屈性 nutritropism)。本研究ではこの伸長方向の変化に植物ホルモンの合成や輸送が関与していることを明らかにしました。
発表概要
東京大学農学生命科学研究科の山崎清志、大森良弘、藤原徹は名古屋大学の研究グループ、国立遺伝学研究所の研究グループの共同研究により、イネの根がアンモニウムの濃度勾配を感知し、その伸長方向をアンモニウム濃度が高い方向に変化させる際、根端では3つの植物ホルモンであるオーキシン、ジベレリン、エチレンのシグナルが重要な働きをしていることを発見しました。
外界の光や重力、水分などの方向性のある刺激に対して、植物が示す方向性のある生長応答を屈性と呼び、その刺激を屈性刺激といいます。屈性刺激を感知した植物は、外界の屈性刺激の分布を体内にある植物ホルモンの分布に変換することが知られており、これを屈性研究ではシグナル変換と呼んでいます。しかしながら、最近イネで発見されたばかりのアンモニウムに対する栄養屈性については、アンモニウムからどのようなシグナル変換が生じるのか不明でした。本研究成果によって、イネの根がアンモニウムの方向を感知したとき、その方向はオーキシン、ジベレリン、エチレンの分布にシグナル変換される必要があることが明らかになりました。
この研究成果は、2023年1月16日に学術誌「Plant and Cell Physiology」にてオンライン公開されました。
この研究は、日本学術振興会(JSPS) 科学研究費補助金の支援を受けて行われました。
発表内容
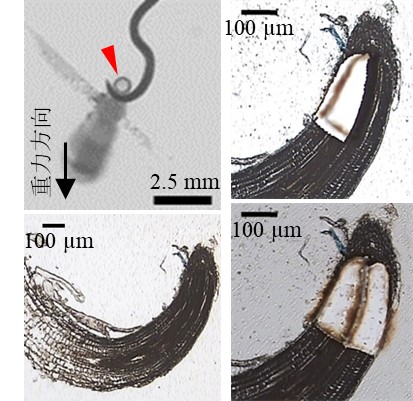
図1:イネ主根における栄養屈性と根端内側および外側組織の切り出し
イネ幼苗の主根の伸長方向にアンモニウムを含む栄養源を設置すると、6時間後には栄養源に巻き付き始める(左上図)。栄養源に巻き付いた根端のパラフィン切片を作成し観察すると、顕微鏡下でも栄養屈性の屈曲方向が観察できる(左下図)。レーザーマイクロダイセクションという手法で、屈曲している根の内側と外側の組織を切り出し回収した(右図)。回収した組織からmRNAを抽出し、次世代シーケンサーを用いたRNA-seq解析によって組織間の遺伝子発現の違いを調査した。

図2:根端内側および外側組織で発現差を示した遺伝子解析
内側組織と外側組織で有意な発現差を示した153遺伝子について、発現パターンをクラスタリングすると内側組織(i1~i6)と外側組織(o1~o6)の2グループに明瞭に分かれた (左図)。発現差を示した遺伝子群からオーキシン、エチレン、ジベレリン、サリチル酸シグナリングに関与するものが検出された(右図)。
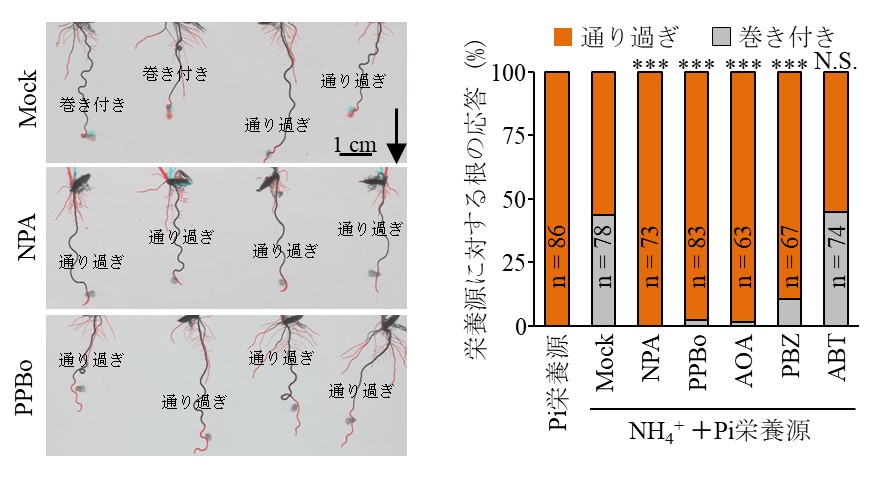
図3:各植物ホルモン輸送または合成阻害剤を栄養源に加えた時の栄養屈性
アンモニウム(NH4+)とリン酸(Pi)を含む栄養源に各阻害剤を添加したものを用いて栄養屈性の試験を実施した。左図は阻害剤なし(Mock)、オーキシン輸送阻害剤(NPA)、オーキシン合成阻害剤(PPBo)を添加したときの例を示す。Mock条件では約半数の個体の根で栄養屈性を示し栄養源に巻き付く。栄養源設置後の12時間で伸長した根は赤で示されている。右図は、各種の阻害剤を添加した栄養源に対して、栄養屈性(巻き付き)を示した個体の割合。栄養屈性を示さない条件として、栄養屈性刺激(アンモニウム)を含まないPi栄養源に対する応答も参考値として計測している。測定の結果、NPA、PPBo、エチレン合成阻害剤(AOA)、及びジベレリン合成阻害剤(PBZ)を添加した条件で有意に栄養屈性が阻害され、サリチル酸合成阻害剤(ABT)添加条件ではMockと同等の応答性を示した。矢印: 重力方向, ***: 統計的に有意(p < 0.001)な差.
研究背景と内容
植物の屈性応答は、屈性刺激の感知、シグナル変換、偏差生長の3つの応答に分類することができます。根の重力屈性と水分屈性では、屈性刺激が感知されると、植物ホルモンであるオーキシン、サイトカイニン応答にそれぞれシグナル変換されます。重力屈性ではオーキシンシグナルの下流で細胞伸長が、水分屈性ではサイトカイニンシグナルの下流で細胞分裂が制御されることにより、根が屈曲するとされています。
最近我々が発見した栄養屈性では、シグナル変換で植物ホルモンが関与するかさえ不明でした。栄養屈性の刺激がシグナル変換されると、細胞内では変換されたシグナルに応じて、屈性刺激側とその反対側で異なる遺伝子発現が生じているはずです。しかし、『栄養屈性を示している最中の根』から『屈性刺激側とその反対側に位置する根端組織を区別して単離』することは難題です。
本研究では、栄養屈性刺激であるアンモニウム栄養源に巻き付いている最中のイネ主根を『栄養屈性を示している最中の根』として取得しました(図1左)。この根から作成した根端切片では、巻き付いている方向から栄養屈性刺激の方向が決定できます。レーザーマイクロダイセクションという技術を用いて、この切片から『屈性刺激側とその反対側に位置する根端組織を区別して単離』し(図1右)、遺伝子発現パターンを比較しました(図2左)。組織間で有意に発現差があった遺伝子のうち、植物ホルモンシグナル伝達に関与する遺伝子を抽出すると、オーキシン、エチレン、ジベレリン、サリチル酸シグナル伝達に関与する遺伝子が検出されました(図2右)。これら植物ホルモンの合成または輸送の阻害剤を、イネの根の伸長を阻害しない濃度で栄養源に添加したところ、オーキシン輸送阻害剤、オーキシン、エチレン、ジベレリン合成阻害剤を添加したとき、栄養屈性は強く阻害され、これらの植物ホルモンシグナル伝達が栄養屈性に必要であることが分かりました(図3)。
本成果で、植物根の栄養屈性におけるシグナル変換に関与する植物ホルモンを世界で初めて明らかにすることができました。このシグナル変換に関与したオーキシン、エチレン、ジベレリンシグナリングは重力屈性でも重要な働きをしていることが知られており、栄養屈性と重力屈性は屈曲のメカニズムが似ているのかもしれません。
成果の意義
栄養屈性におけるシグナル変換に関与する植物ホルモンの特定により、栄養屈性が重力屈性と同様のホルモンシグナル伝達を介して屈曲していることが分かりました。このことから、栄養屈性刺激の方向に依存した植物ホルモンの分布は、重力屈性刺激にも影響を受けていると考えられ、植物の根が栄養屈性を示すか重力屈性を示すかは、ホルモン分布の競合というモデルで説明できます。今後、本研究で組織間に発現差があった遺伝子から、実際に栄養屈性に関与する遺伝子が同定されることで、さらなる分子機構の解明が期待されます。
発表雑誌
- 雑誌名
- Plant and Cell Physiology
- 論文タイトル
- Transcriptome analysis of rice root tips reveals auxin, gibberellin, and ethylene signalling underlying nutritropism
- 著者
- Kiyoshi Yamazaki*, Yoshihiro Ohmori, Hirokazu Takahashi, Atsushi Toyoda, Yutaka Sato, Mikio Nakazono, Toru Fujiwara*
- DOI番号
- 10.1093/pcp/pcae003
- 論文URL
- https://academic.oup.com/pcp/advance-article/doi/10.1093/pcp/pcae003/7558230
問い合わせ先
東京大学大学院農学生命科学研究科 応用生命化学専攻 植物栄養・肥料学研究室
教授 藤原 徹 (ふじわら とおる)
Tel/Fax: 03-5841-5104
E-mail: atorufu[アット] g.ecc.u-tokyo.ac.jp
東京大学大学院農学生命科学研究科 応用生命化学専攻 植物栄養・肥料学研究室
特任講師 山崎 清志 (やまざき きよし)
Tel/Fax: 03-5841-5104
E-mail: aykiyo[アット] g.ecc.u-tokyo.ac.jp [アット]を@に変えてください。