「強光でも負けない植物!」生産性向上の鍵となる画期的な光合成促進剤を発見! ――過酷な環境での収穫量向上に貢献する植物成長調整剤の開発へ期待――
発表のポイント
- 地球温暖化や異常気象が引き起こす強い日差し(強光ストレス)は、植物の成長を妨げ、作物の収穫量を大幅に減少させる深刻な課題です。
- 本研究では、アントラキノン(3つのベンゼン環が繋がった構造)の仲間が、強い日差しの下で植物の光合成能力を高め、植物を健康に育てることを発見しました。この物質はタバコ、レタス、トマトなどの多様な植物に効果を発揮し、葉にスプレーするだけでストレス環境下での植物成長を促進することが明らかになりました。
- 本技術の活用により、過酷な環境でも作物を安定して育てることが可能となり、世界の食料問題解決に大きく貢献することが期待されます。
図1:「強光ストレスを克服!」植物の生産性を劇的に高める光合成促進剤の発見
研究概要
東京大学大学院農学生命科学研究科の矢守航准教授らは、強い日差しにも負けず、植物の生産性を高める画期的な光合成促進剤を発見しました。
地球温暖化や異常気象によって、強い日差し(強光ストレス)が植物の成長を妨げ、作物の収穫量を大幅に減らしてしまうことが大きな課題となっています。増え続ける世界人口を支えるため、食料生産を2050年までに現在より50%増やす必要があるとされていますが、このような環境ストレスがその実現を阻む大きな障害となっています。
本研究グループはこの課題を解決するため、植物を守る新しい化合物を探す独自のスクリーニングシステムを開発しました。このシステムを使って発見されたのが「アントラキノン(3つのベンゼン環が繋がった構造)」という化合物の仲間です。この化合物を葉にスプレーすることで、タバコ、レタス、トマトなどの植物が強い日差しの中でも光合成能力を保ち、元気に成長できることが確認されました。
さらに、この化合物は、植物がストレスを受けていない普通の環境でも悪影響を及ぼさないことも明らかになりました。これらにより、過酷な環境でも安定した作物生産が可能となり、世界的な食料問題の解決に向けた大きな一歩となることが期待されます。
研究内容
2050年までに世界人口は100億人に達すると予測され、その人口を支えるためには現在の食料生産を50%増やす必要があります。しかし、干ばつや洪水、強い日差し(強光ストレス)など、気候変動による自然災害が農業生産に深刻な影響を及ぼしています。特に、強光ストレスによる光合成能力の低下は作物の成長と収量を阻害する大きな要因です。これまで、ゲノム編集や形質転換技術によって植物の機能を強化する取り組みが進められてきましたが、食料生産性を向上させるためには、化合物を利用したストレス耐性向上技術など、多角的なアプローチが必要とされています。
東京大学大学院農学生命科学研究科の曲玉辰(Qu Yuchen)博士研究員と矢守航准教授らは、植物が強光ストレス下でも効率的に光合成を行い、健康に成長できる化合物を探し出すための独自のスクリーニングシステムを開発しました。このシステムを活用して、12,000種類の化合物を光合成の可視化装置で解析したところ、「アントラキノン(3つのベンゼン環が繋がった構造)」と呼ばれる化合物の仲間が強光ストレス環境下で光合成を促進することを発見しました(図1, 2)。
光合成の過程では、光化学系II(PSII)と光化学系I(PSI)が連携し、電子伝達を通じて光エネルギーをATPやNADPHに変換します。そこで次に、アントラキノンの仲間(A1NとA4N)が光合成の電子伝達過程において電子受容体として機能する可能性を調べました(図3)。その結果、これらの化合物はPSIIでは電子を受け取らない一方、PSIからは電子を部分的に受け取る能力を持つことを確認しました(図3)。
また、タバコの葉におけるアントラキノンの仲間(A1N)の効果を解析しました。強い光ストレスを72時間与えた後、タバコの葉では光合成効率やクロロフィル濃度が低下しましたが、アントラキノンの仲間(A1N)を葉にスプレーしたグループではこれらの低下が顕著ではなく、葉の緑色も保たれていました(図4A)。一方、アントラキノン類(A1N)処理の有無にかかわらず、二酸化炭素の取り込みに関わる気孔開度に差は見られませんでした。さらに、光ストレスが光合成の仕組みに与える影響を詳しく調べたところ、光化学系II(PSII)と光化学系I(PSI)の電子伝達活性(ETR I、ETR II)は、アントラキノンの仲間(A1N)を処理したグループで顕著に高いことが分かりました(図4B)。特に強い光の条件下では、アントラキノンの仲間(A1N)が光化学系Iの効率(Y(I))を向上させ、光エネルギーを無駄なく利用していることが分かりました(図4B)。同様の結果を、シロイヌナズナ、レタス、トマトにおいて確認しました。
さらに、強い日差し(光ストレス)を受けた後の植物の回復や成長に、アントラキノンの仲間(A1N)がどのような効果を与えるかを解析しました。シロイヌナズナ(図5)、レタス(図6)、トマト(図7)を用いて実験を行ったところ、アントラキノンの仲間(A1N)を処理した植物では、光ストレスを受けた後も光化学系IIの光合成効率(Y(II))が高く保たれ、光エネルギーの無駄(Y(NPQ)やY(NO))が抑えられることが分かりました。トマトでは、アントラキノン類(A1N)処理により光ストレス後のクロロフィル量や光合成装置の健全性(Fv/Fm)が向上しました(図7)。さらに、シロイヌナズナを対象に網羅的遺伝子発現解析を行ったところ、光合成や炭素固定に関わる遺伝子群がアントラキノンの仲間(A1N)処理によってより活発に機能していることを明らかにしました(詳細は論文をご覧ください)。光ストレス後に通常の環境で1週間回復させた際、アントラキノンの仲間(A1N)を処理した植物は成長が良好で、ストレスの影響がほとんど見られませんでした(図5C、6C、7C)。一方、未処理の植物では成長が抑制され、早い開花や葉の変色が観察されました(図5C、6C、7C)。これらの結果は、アントラキノンの仲間(A1N)が植物の光ストレス耐性を高め、回復を助ける可能性を示します。
さらに重要なのは、アントラキノンの仲間(A1N)を処理した植物が通常の環境下(ストレスのない条件)では、特に悪影響を受けなかった点です。具体的には、アントラキノンの仲間(A1N)を処理したタバコの葉では、通常の栽培環境で継続して育成された場合、光合成装置の健全性(Fv/Fm)においてコントロール群(未処理の植物)とほとんど差が見られませんでした。
アントラキノンの仲間は自然界に広く存在し、植物の花や果実、根茎に含まれる化合物です。これらの物質はUV-B保護やエネルギー伝達調整、抗炎症作用などで知られ、医療や農業分野で幅広く利用されています。本研究では、アントラキノンの仲間がタバコ、レタス、トマト、シロイヌナズナで光ストレスを軽減し、光合成能力や植物成長を助けることを確認しました。さらに、この化合物はストレス環境での安全性も高く、通常の栽培条件では7日以内に分解されることもわかっています。アントラキノンの仲間は、農業における光ストレス耐性向上剤として、作物の生産性向上や高価値作物(果物、野菜、花卉など)の品質維持に貢献する可能性があります。この研究成果は、気候変動による農業課題を克服し、持続可能な食料供給の実現に向けた新しい技術として期待されます。
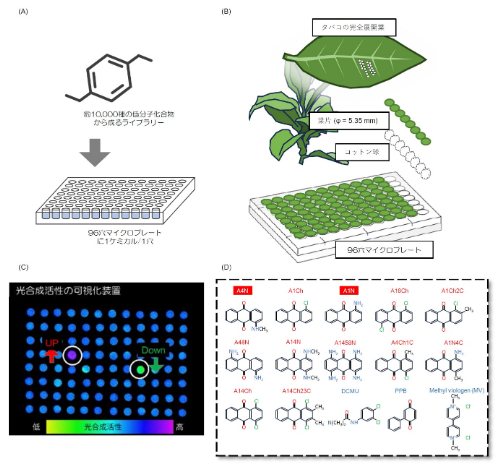
図2:タバコのリーフディスクを使用したケミカルスクリーニングシステムの構築
(A) 96ウェルプレートを利用した化合物ライブラリー。(B) 綿球を入れた96ウェルプレートの各穴に各化合物を添加し、その上にリーフディスクを静置した。(C) 化合物を処理したリーフディスクの光合成活性を、IMAGING-PAMシステムを用いて可視化した。(D) 光ストレスに対して保護効果を示した候補化合物(アントラキノン類;赤文字)と、実験で使用した3つの阻害剤/電子受容体(青文字)の構造式。DCMU; ジウロン、PPB; フェニル-p-ベンゾキノン、MV; メチルビオローゲン
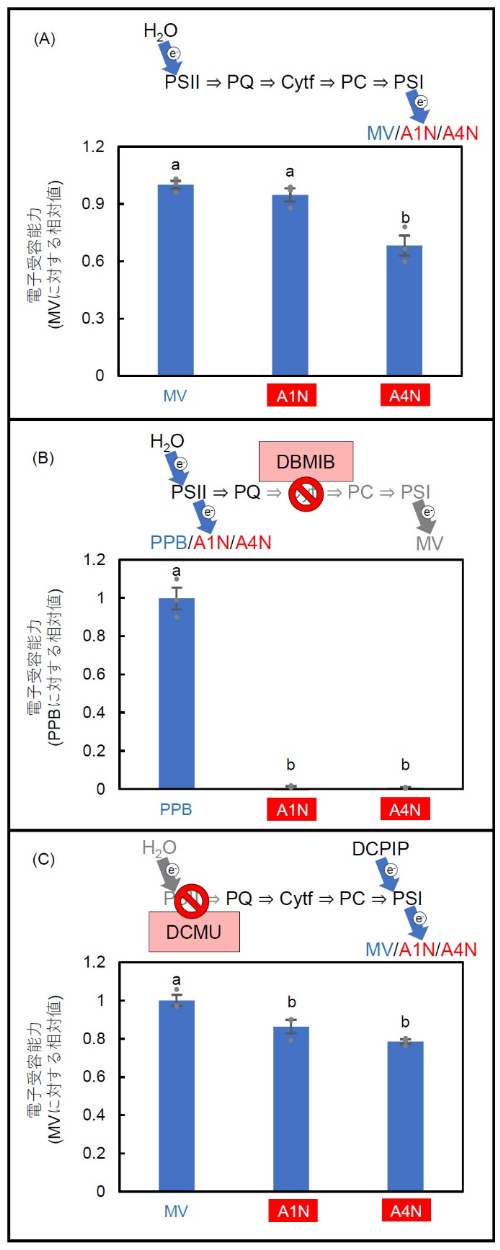
図3:光合成電子伝達系におけるアントラキノン類(A1N、A4N)の電子受容能力
(A) 光合成電子伝達系におけるアントラキノン類の電子受容能力をMV(メチルビオローゲン)と比較した。各数値はMVの電子受容能力を1としたときの相対値。(B) 光化学系II(PSII)におけるアントラキノン類の電子受容能力をPPB(フェニル-p-ベンゾキノン)と比較した。各数値はPPBの電子受容能力を1としたときの相対値。(C) 光化学系I(PSI)におけるアントラキノン類の電子受容能力をMVと比較した。各数値はMVの電子受容能力を1としたときの相対値。各測定により予想された各化合物の作用機序をグラフの上部に示す。各数値は平均値 ± 標準誤差(SE)で示され、異なる英文字間には5%水準で有意差があることを示す(n = 4)。
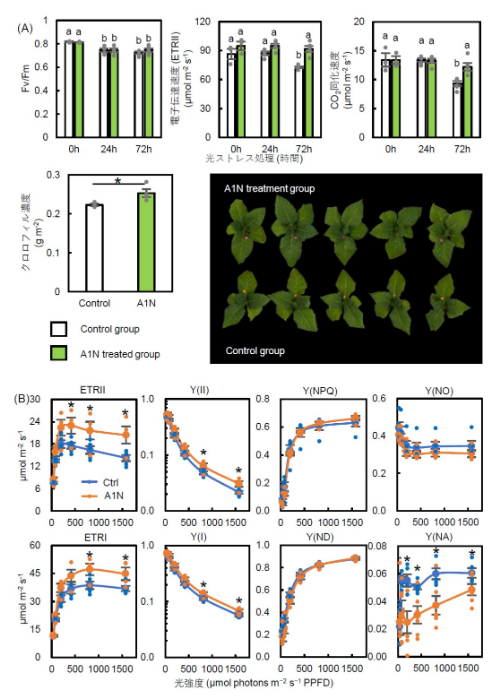
図4:光ストレス処理後のタバコの葉の光合成能力
(A) 光ストレス処理0時間、24時間、72時間後の光化学系II(PSII)の最大量子収率(Fv/Fm)、PSIIの電子伝達速度(ETRII)、およびCO2同化速度。光ストレス処理72時間後のクロロフィルa+b濃度とタバコ植物の写真。アントラキノン類(A1N)処理グループの葉はピンクのステッカー、コントロールグループ(Ctrl)の葉は黄色のステッカーでマークした。(B) 光ストレス処理72時間後における光化学系II(PSII)と光化学系Iの光合成パラメータ*1の光応答。各数値は平均値 ± 標準誤差(SE)で示され、異なる英文字間および*は5%水準で有意差があることを示す。
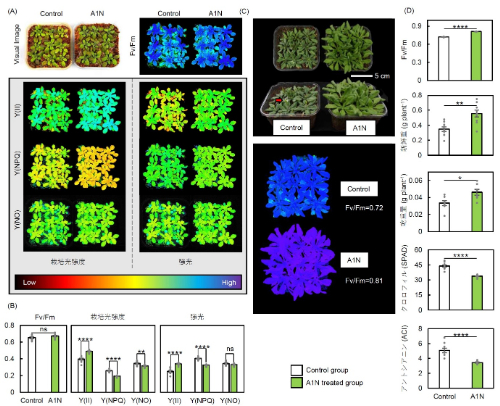
図5:光ストレス処理96時間後と処理後1週間回復させたシロイヌナズナの光合成能力
(A, B) 光ストレス処理96時間後における光ストレス条件(700 μmol photons m⁻² s⁻¹)と栽培光条件(200 μmol photons m⁻² s⁻¹)での光化学系II(PSII)の光合成パラメータ*1の可視化画像と測定値(Fv/Fm、Y(II)、Y(NPQ)、Y(NO))。(C) 回復1週間後における植物体の画像とFv/Fmの可視化画像。赤い矢印は早期の開花を示す。(D) 回復1週間後におけるFv/Fm、地上部の新鮮重量、地上部の乾物重量、SPAD(クロロフィル含有量)およびアントシアニン含有量。アントラキノン類(A1N)処理ありと処理無し(Control)で比較した。各数値は平均値 ± 標準誤差(SE)で示され、*、**、***、****は各々5%、1%、0.1%、0.01%水準で有意差があること、nsは有意差がないことを示す (n = 9) 。
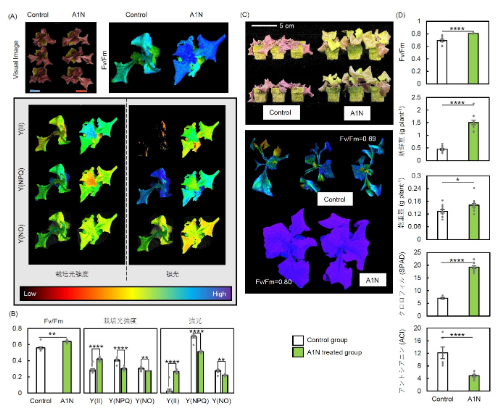
図6:光ストレス処理96時間後と処理後1週間回復させたレタスの光合成能力
(A, B) 光ストレス処理96時間後における光ストレス条件(700 μmol photons m⁻² s⁻¹)と栽培光条件(200 μmol photons m⁻² s⁻¹)での光化学系II(PSII)の光合成パラメータ*1の可視化画像と測定値(Fv/Fm、Y(II)、Y(NPQ)、Y(NO))。(C) 回復1週間後における植物体の画像とFv/Fmの可視化画像。(D) 回復1週間後におけるFv/Fm、地上部の新鮮重量、地上部の乾物重量、SPAD(クロロフィル含有量)およびアントシアニン含有量。アントラキノン類(A1N)処理ありと処理無し(Control)で比較した。各数値は平均値 ± 標準誤差(SE)で示され、*、**、***、****は各々5%、1%、0.1%、0.01%水準で有意差があることを示す (n = 9) 。
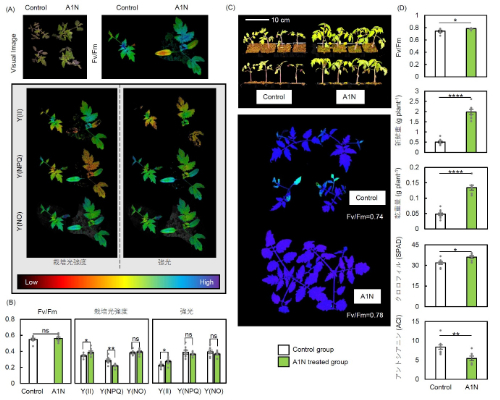
図7:光ストレス処理96時間後と処理後1週間回復させたトマトの光合成能力
(A, B) 光ストレス処理96時間後における光ストレス条件(700 μmol photons m⁻² s⁻¹)と栽培光条件(200 μmol photons m⁻² s⁻¹)での光化学系II(PSII)の光合成パラメータ*1の可視化画像と測定値(Fv/Fm、Y(II)、Y(NPQ)、Y(NO))。(C) 回復1週間後における植物体の画像とFv/Fmの可視化画像。(D) 回復1週間後におけるFv/Fm、地上部の新鮮重量、地上部の乾物重量、SPAD(クロロフィル含有量)およびアントシアニン含有量。アントラキノン類(A1N)処理ありと処理無し(Control)で比較した。各数値は平均値 ± 標準誤差(SE)で示され、*、**、****は各々5%、1%、0.01%水準で有意差があること、nsは有意差がないことを示す (n = 9) 。
※各実験のパラメータ詳細
〇クロロフィル蛍光を用いた光化学系II(PSII)のパラメータ
・Fv/Fm:PSIIの潜在的な最大効率とその健全性の指標
・Y(II):PSIIで光エネルギーが有効に利用される割合
・Y(NPQ):植物が過剰な光エネルギーを熱として散逸させる割合
・Y(NO):光によって誘導されないエネルギー散逸(熱放散や損傷を含む)の割合
・ETRII:PSIIにおける光合成電子伝達速度の指標
・qP:プラストキノンの酸化還元状態の指標
*Y(II)+Y(NPQ)+Y(NO)=1となる。
〇P700吸収変化を用いた光化学系I(PSI)のパラメータ
・Pm:PSIの酸化能力とその健全性の指標
・Y(I):PSIで光エネルギーが有効に利用される割合
・Y(NA):PSIの電子受容体が制限要因となる割合(高い値は、カルビン回路や電子受容体(フェレドキシンやNADP+)に制約があることを示す。)
・Y(ND):PSIの電子供給側が制限要因となる割合(高い値は、PSIIやシトクロムb6/f複合体からPSIへの電子供給が制約されていることを示す。)
・ETRI:PSIにおける光合成電子伝達速度の指標
*Y(I)+Y(ND)+YNA)=1となる。
発表者
東京大学
大学院農学生命科学研究科
曲 玉辰(Qu Yuchen) 博士研究員
迫田 和馬 研究当時:博士研究員
若林 侑 助教
中嶋 正敏 教授
浅見 忠男 東京大学名誉教授
矢守 航 准教授
大学院理学系研究科
寺島 一郎 東京大学名誉教授
論文情報
- 雑誌
- Communications Biology
- 題名
- Identification and characterization of compounds that improve plant photosynthesis and growth under light stress conditions
- 著者
- Qu Yuchen, Sakoda Kazuma, Wakabayashi Yu, Nakajima Masatoshi, Asami Tadao, Terashima Ichiro, Yamori Wataru *
- DOI
- 10.1038/s42003-025-07582-2
研究助成
本研究は、日本学術振興会科研費「基盤研究(S)(課題番号:20H05687)」、「基盤研究(B)(課題番号:21H02171)」、「学術変革領域研究(A)「細胞質ゲノム制御」(課題番号:24H02277、24H02270)」の支援により実施されました。
問い合わせ先
(研究内容については発表者にお問合せください)
東京大学大学院農学生命科学研究科附属生態調和農学機構
准教授 矢守 航(やもり わたる)
TEL: 070-6442-9511 E-mail: yamori[at]g.ecc.u-tokyo.ac.jp
東京大学大学院農学生命科学研究科・農学部
事務部 総務課総務チーム 総務・広報情報担当(広報情報担当)
TEL: 03-5841-8179, 5484 FAX:03-5841-5028 E-mail: koho.a[at]gs.mail.u-tokyo.ac.jp
※上記の[at]は@に置き換えてください。