葉化病の病原タンパク質「ファイロジェン」の機能は1アミノ酸の変異で制御される ―水平移動による進化と病原性の変異メカニズムを解明―
- 発表者
- 岩渕 望 (東京大学大学院農学生命科学研究科 生産・環境生物学専攻 特任研究員)
北沢 優悟 (東京大学大学院農学生命科学研究科 生産・環境生物学専攻 特任研究員)
前島 健作 (東京大学大学院農学生命科学研究科 生産・環境生物学専攻 助教)
発表のポイント
- ファイトプラズマの葉化誘導因子「ファイロジェン」が、水平移動による独自の進化を遂げたことを解明しました。
- ファイロジェンの機能比較により、わずか1アミノ酸の変異がファイトプラズマの病原性を決定していることが初めて明らかになりました。
- 本成果をもとにした、ファイトプラズマの病原性の進化機構の解明ならびに葉化病の治療薬の開発が期待されます。
発表概要
植物病原細菌ファイトプラズマ(注1)は、様々な植物に葉化(注2、図1)や天狗巣(注3)など特徴的な形態異常を引き起こします。ファイトプラズマが持つ葉化誘導因子ファイロジェンは、植物の花形成に重要な因子と類似した構造を持つことでその働きを妨げ、葉化を誘導します。しかし、それぞれのファイトプラズマがもつファイロジェンの機能の保存性、また葉化を引き起こすのに重要なアミノ酸部位は不明でした。
今回、東京大学大学院農学生命科学研究科の岩渕望特任研究員と前島健作助教らは、新しく開発したファイロジェンの探索法により、多様なファイトプラズマからファイロジェンを網羅的に特定することに成功し、ファイロジェンがファイトプラズマ間を水平移動することによりファイトプラズマとは独立に進化していることを明らかにしました。さらに、新しく同定したファイロジェンの中から、進化の過程で葉化を引き起こさなくなったファイロジェン(非葉化型ファイロジェン)の一群を初めて発見すると共に、その原因がわずか1アミノ酸の変異により説明できることを明らかにしました。
本成果は、ファイトプラズマの病原性がどのように決定されているのかを明らかにした初めての成果です。
発表内容

図1 ファイトプラズマの感染により花が葉化したアサガオ
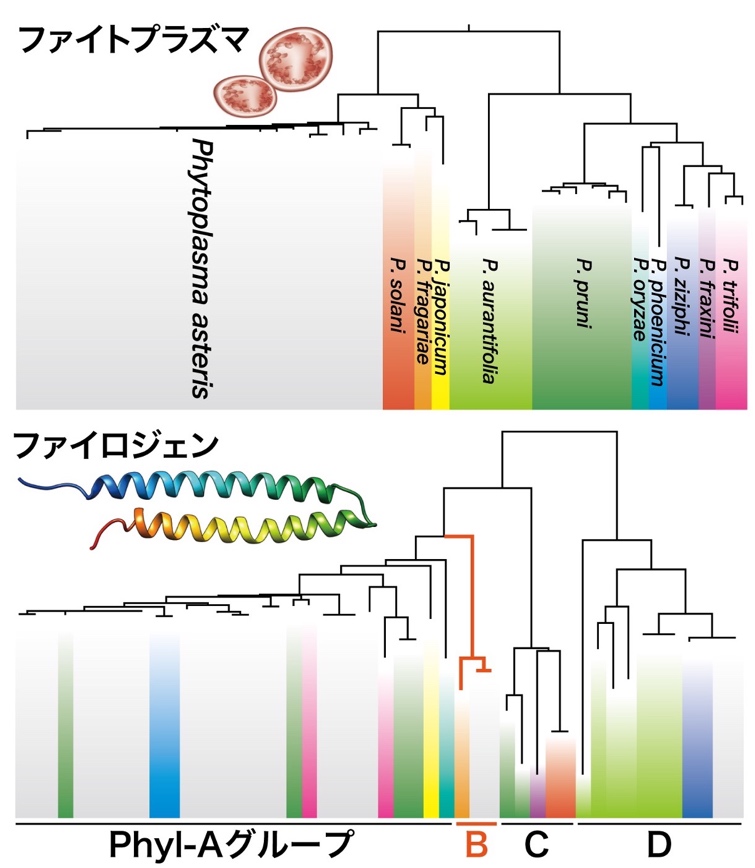
図2 ファイロジェンは水平移動による独自の進化を遂げていた
系統樹の背景の色はそれぞれのファイトプラズマが属する種を表す。遠縁なファイトプラズマが非常によく似たファイロジェンをもつ場合があるなど、ファイトプラズマの進化(上段)に対して葉化誘導因子「ファイロジェン」の進化(下段)は独立に起きていた。これは、ファイロジェンがファイトプラズマ間を水平移動しているためと考えられた。また、ファイロジェンが進化の過程で4グループに分かれ、Phyl-Bグループ(オレンジ色)は葉化を引き起こさなくなったことが明らかになった。
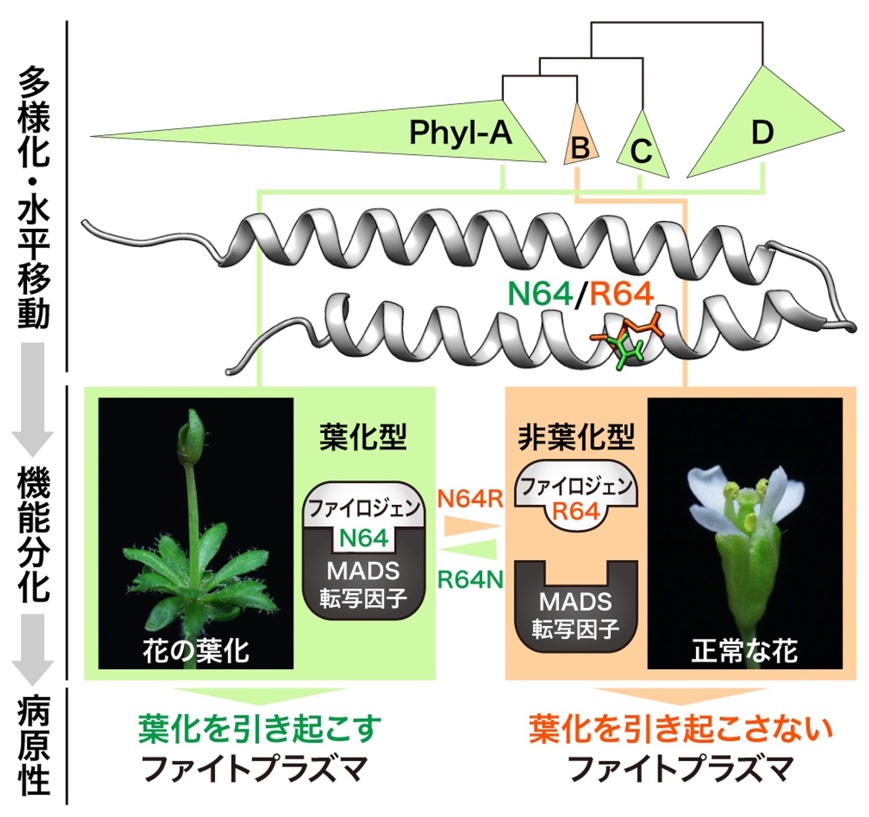
図3 ファイロジェンの1アミノ酸の違いによる病原性決定のモデル図
Phyl-Bグループに特有の1アミノ酸変異(64番目のアミノ酸がアスパラギン[N64: 緑色]からアルギニン[R64: オレンジ色]に変異)が葉化を引き起こすかどうかを決定していた。N64をもつファイロジェンは植物のMADS転写因子への結合・分解活性をもつ葉化型になり、R64をもつファイロジェンは活性のない非葉化型になった。すなわちこのわずか1アミノ酸の違いによりファイトプラズマの病原性が決定されていることが明らかになった。
ファイトプラズマは1,000種以上の植物の篩部に寄生し、ヨコバイ等の昆虫により媒介される微小な植物病原細菌で、世界中の農業生産に甚大な被害をもたらしています。病原細菌は多種多様な病原性因子を用いて、宿主の細胞機能を自身の感染成立に有利になるように改変することが知られています。中でもファイトプラズマからは、植物の花を葉に変化させるファイロジェン(注4)や、小さな枝葉を無数に生じさせるTENGUなど、植物のダイナミックな形態変化を引き起こすユニークな病原性因子が見いだされています。
花器官の形成にはきわめて複雑な遺伝子の発現制御が必要ですが、その制御を司っているのが植物に共通して存在する花形成因子(MADS転写因子[注5])の多量体化です。東京大学農学部の植物病理学研究室では、昨年、世界で初めてファイロジェンの立体構造(注6)を解明し、ファイロジェンがMADS転写因子の多量体化領域と類似した立体構造を有することで、MADS転写因子を標的とした結合・分解を引き起こすことを明らかにしました。ファイロジェンはその立体構造を壊すと機能を失いますが、ファイトプラズマ間での立体構造の保存性や、立体構造の中で葉化誘導に重要なアミノ酸部位はこれまで明らかになっていませんでした。
今回、同研究室の岩渕望特任研究員と前島健作助教らは、新しく開発したファイロジェンの網羅的探索法により、ファイトプラズマの多様な種や系統からファイロジェンの特定に成功しました。その結果、ファイトプラズマ間でファイロジェンの立体構造は保存されているものの、これまで想定されていた以上に配列多様性に富み、ファイトプラズマとは独立に進化していることが示されました(図2)。その原因として、ファイトプラズマ間でファイロジェン遺伝子の水平移動(注7)が起きていたことが明らかになりました。また、機能の比較解析により、ファイロジェンが進化の過程で、葉化を引き起こす「葉化型」と引き起こさない「非葉化型」のグループに機能分化していることを明らかにしました(図3)。さらに、非葉化型に特徴的なアミノ酸残基の置換解析により、立体構造の中で64番目のアミノ酸がMADS転写因子への結合・分解と葉化を引き起こすかどうかを決定していることを明らかにしました(図3)。
本研究成果は、ファイトプラズマによる葉化病がファイロジェンという単一のタンパク質の、わずか1アミノ酸の変異によって決定されていることを示しています。本研究により、ファイトプラズマの病原性の進化と変異の仕組みが初めて解明されました。
ファイトプラズマは栄養分を全て宿主から得ているため、自身は代謝に関わる遺伝子の大半を失う退行的進化を遂げています。一方で、今回の研究により、ファイロジェン遺伝子に関しては水平移動を伴う積極的な獲得が起きていることが示されました。このことは、ファイロジェン遺伝子がファイトプラズマの生存を有利にする重要な働きを担っていることを示しています。
興味深いことに、ファイロジェンは葉化誘導に重要なMADS転写因子以外にも、様々な植物因子と相互作用するほか、媒介昆虫の誘引など多面的機能をもつことが示唆されています。今回特定した非葉化型ファイロジェンも、MADS転写因子への結合能の喪失とひきかえに、新たな植物因子を標的とする未知の機能を獲得している可能性があります。今後、ファイロジェンの多様性とその分子機能の全体像を解明することで、ファイトプラズマの感染メカニズムの解明や、ファイロジェンの機能部位を標的とした治療薬の開発につながることが期待されます。
本研究は日本学術振興会科学研究費補助金の支援を受けて行われました。
発表雑誌
- 雑誌名
- Molecular Plant Pathology(8月19日オンライン公開)
- 論文タイトル
- Functional variation in phyllogen, a phyllody-inducing phytoplasma effector family, attributable to a single amino acid polymorphism
- 著者
- Nozomu Iwabuchi, Yugo Kitazawa, Kensaku Maejima, Hiroaki Koinuma, Akio Miyazaki, Ouki Matsumoto, Takumi Suzuki, Takamichi Nijo, Kenro Oshima, Shigetou Namba and Yasuyuki Yamaji
- DOI番号
- 10.1111/mpp.12981
- 論文URL
- https://bsppjournals.onlinelibrary.wiley.com/doi/abs/10.1111/mpp.12981
問い合わせ先
東京大学大学院農学生命科学研究科 生産・環境生物学専攻 植物病理学研究室
助教 前島 健作 (まえじま けんさく)
Tel:03-5841-1613
Fax:03-5841-5090
E-mail:amaejima<アット>mail.ecc.u-tokyo.ac.jp <アット>を@に変えてください。
研究室URL:http://park.itc.u-tokyo.ac.jp/ae-b/planpath/
用語解説
- 注1 ファイトプラズマ
東京大学農学部の植物病理学研究室において、1967年にマイコプラズマ様微生物(mycoplasma-like organism, MLO)として発見され、2004年に全ゲノムが解読された、ファイトプラズマ属(モリキューテス綱)に分類される植物病原細菌。細胞壁を欠いた直径0.1〜0.8 μmの不ぞろいな粒子状で、細菌の中でも最小サイズ。1,000種以上の植物に感染し、世界中の農業生産に被害をもたらしている。植物では篩部にのみ寄生し、ヨコバイなどの篩部吸汁性昆虫により媒介される。植物に萎縮病、天狗巣病、葉化病(花の葉化・緑化・突き抜け)などの特徴的な病気を引き起こし、植物を枯らしてしまうことも多い。ファイトプラズマ病は、私たちの身近にも頻繁に認められる病気であり、葉化病によって緑色の花が咲くアジサイは商品価値が認められ、品種登録されていた例もある。また、クリスマスシーズンの風物詩である鉢植えのポインセチア(枝分れが豊富で、矮性化するタイプ)は、全て背丈を小さくするためにファイトプラズマに人工的に感染させられ、天狗巣病を発病したものであり、健全なポインセチアは数メートルの高さにもなる。 - 注2 葉化
花びらやがく・雌しべ・雄しべが葉に置き換わる症状。 - 注3 天狗巣
側芽が異常に発達し、小枝が密生する症状。 - 注4 葉化誘導因子 ファイロジェン
「葉化病(phyllody、phyllo-(葉)+-ody(になる変化))」の原因である遺伝子ファミリー「phyllody-inducing gene family(葉化誘導遺伝子ファミリー)」の略。phyllo-(葉)+-gen(を生ずるもの=gene(遺伝子))。ファイロジェンはあらゆる植物の花形成因子を標的とすることで、全ての葉化病をファイロジェン1つで引き起こすことができると考えられる。(以下、関連のプレスリリース)
・「花」を「葉」に変える病気の謎を解く ―原因遺伝子の発見と発症メカニズムの解明―【2014年3月18日】
・1つの病原性因子があらゆる植物種の花形成因子を分解する ―葉化病発症メカニズムは植物に共通であることを解明―【2017年5月16日】 - 注5 MADS転写因子
花の形成を制御する転写因子。どの植物細胞が花びらやがくになるかは、MADS転写因子の組み合わせによって決まる。4つのMADS転写因子は多量体(四量体)を形成して機能すると考えられており、多量体形成にはMADS転写因子を構成する4つのドメインのうち、Kドメインが関わる。MADS転写因子どうしの多量体化の分子機構は植物間で高度に保存されている。 - 注6 ファイロジェンの立体構造
ファイロジェンは約90アミノ酸程度の小さなタンパク質で、MADS転写因子どうしの多量体化に重要なKドメインと類似した立体構造を持つ。これにより、ファイロジェンはその分子機構を模倣し、あらゆる植物のMADS転写因子を標的にすると考えられる。実際に、ファイロジェンの立体構造を壊すような変異を入れると、MADS転写因子に対する結合・分解活性が失われるとともに、植物への葉化誘導能が失われる。(以下、関連のプレスリリース)
・葉化病発症の仕組みを構造化学的に解明 ―葉化病治療薬開発や新品種開発に新たな道―【2019年4月19日】 - 注7 遺伝子の水平移動
生物は、生息域が同じ生物や環境中から遺伝子を自身のゲノムに取り込む場合がある。このようにして、個体間や他の生物間で生じる遺伝子のやりとりを「遺伝子の水平移動」と呼ぶ。毒素を産生する病原菌の出現や、抗生物質が効かなくなる耐性菌の出現は、微生物間での遺伝子の水平移動が一因と考えられる。