細菌胞子の休眠と覚醒の制御に関する新たな知見:放線菌の胞子嚢胞子は完全には休眠していない?
- 発表者
- 手塚 武揚 (東京大学大学院農学生命科学研究科 応用生命工学専攻 農学共同研究員(元同専攻助教)、北里大学 大村智記念研究所・感染制御科学府 特任助教)
光山 京太 (東京大学大学院農学生命科学研究科 応用生命工学専攻 修士課程(当時))
伊達(安田)理沙 (東京大学大学院農学生命科学研究科 応用生命工学専攻 修士課程(当時))
大西 康夫 (東京大学大学院農学生命科学研究科 応用生命工学専攻 教授)
発表のポイント
- 胞子嚢を作る放線菌において、胞子の覚醒の第一段階である胞子嚢開裂の分子機構の解明を目指した研究から、新規な制御様式を有するシグマ/アンチシグマ因子システム(特定の遺伝子群の発現を誘導する仕組みの1つ)の存在が明らかになりました。
- さらに、一連の研究から、胞子嚢開裂の開始を遅らせる分子機構の存在と、それに関与する制御遺伝子が明らかになりました。
- 細菌が形成する胞子は、代謝が完全に停止した休眠状態の細胞であると考えられてきましたが、放線菌の胞子嚢胞子では細胞の休眠状態を維持するために、一部の遺伝子の発現が続いていることが示唆されました。
発表概要
東京大学大学院農学生命科学研究科の手塚武揚博士(元助教、現農学共同研究員)、大西康夫教授の研究グループは、希少放線菌の形態分化の分子機構を長年研究しています(参考URL 1、2)。今回、希少放線菌が形成する胞子嚢(注1)胞子の覚醒の第一段階である胞子嚢開裂の分子機構の解明を目指した研究から、微生物学における新しい概念の提唱につながる重要な成果が得られました。第一に、これまでに知られていないドメイン構成・制御様式をもつシグマ/アンチシグマ因子システム(特定の遺伝子群の発現を誘導する仕組みの1つ)が見出されました。このシグマ/アンチシグマ因子システムは遊走子(注2)の酸化ストレス耐性に関与していることが明らかにされましたが、胞子嚢胞子に特異的な生理学的性質に関与していると推測されます。第二に、本菌は胞子嚢開裂の開始を抑制する仕組みをもっており、それに関与する制御遺伝子が見出されました。この制御遺伝子を過剰発現すると胞子嚢は開裂しなくなる一方、遺伝子破壊により、胞子嚢開裂が野生株より早く開始されるようになりました。以上の結果から、この希少放線菌が作る胞子は、代謝が完全に停止した休眠状態ではなく、休眠状態を維持するために一部の遺伝子の発現を維持している状態であると推測されました。つまり、この放線菌の胞子は、何十年も完全な休眠状態を維持している「植物の種子」というよりは、短期間、活動レベルを低く抑えて生命を維持する「動物の冬眠」に近い性質をもっているのではないかと思われます。細胞レベルで生きている微生物の休眠について、その性質が微生物種によって異なることを示した本研究は、「胞子の生物学」において大きな意義を有しています。
*本成果は2023年12月20日にNature Communications誌にてオンライン公開となりました。
参考URL
(1)注目を集める希少放線菌の不思議に迫る研究(東京大学大学院農学生命科学研究科 研究成果 2017/06/08)
https://www.a.u-tokyo.ac.jp/topics/2017/20170608-1.html
(2)注目を集める希少放線菌の不思議に迫る研究:遊走子線毛の発見と機能解明(東京大学大学院農学生命科学研究科 研究成果 2019/07/10)
https://www.a.u-tokyo.ac.jp/topics/topics_20190710-1.html
発表内容
研究の背景
一部の細菌は、貧栄養など自身の生育に適さない環境におかれると胞子を形成します。胞子は細胞内の代謝活性が低下し、栄養細胞よりも高いストレス耐性を示す休眠耐久状態の細胞として知られており、胞子の形成は自然界における細菌の重要な生存戦略の1つです。一方、病原性細菌の宿主への感染や食品の微生物汚染において、休眠耐久細胞である胞子の存在が感染予防や微生物汚染の防御を困難にしています。そのため、細菌における胞子の形成と覚醒の分子機構の解明は、基礎生物学のみならず病原性細菌の感染予防や食品の微生物汚染の防除という応用面においても、非常に重要な研究領域です。
希少放線菌アクチノプラネス・ミズーリエンシス(Actinoplanes missouriensis)は、菌糸細胞の伸長・分岐により増殖しますが、貧栄養な環境では胞子嚢を形成して休眠状態になります。胞子嚢内部の胞子は、高温や乾燥状態に対して菌糸細胞よりも高い耐性を持ち、長期間の生存を可能にします。一方で、胞子嚢は水がかかると表層の膜(胞子嚢膜)が破れ、内部の胞子を放出します(この過程を胞子嚢の開裂と言います)。胞子嚢から外部に放出された胞子は遊走子となり、増殖に適した環境に到達すると発芽して菌糸細胞の伸長を開始します(図1)。このように、希少放線菌アクチノプラネス・ミズーリエンシスは、細菌としては極めて高度な形態分化能(異なった形態・能力を有する細胞を生じる能力)を有していますが、胞子嚢の開裂は休眠状態の細胞(胞子)が覚醒する最初のステップであるとみなすことができます(図2)。
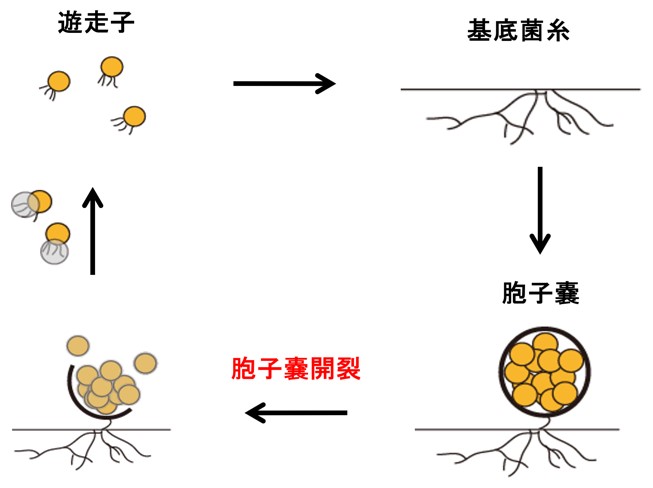
図1. 希少放線菌アクチノプラネス・ミズーリエンシスの生活環
栄養分が枯渇した状態になると、栄養増殖細胞である基底菌糸から、数百の胞子を内包した胞子嚢が形成される。胞子は胞子嚢内部で休眠状態を続けるが、胞子嚢が水に浸されると胞子が休眠から覚醒する。覚醒の第一段階として、胞子嚢の膜が破れ、胞子が水中に放出される(胞子嚢開裂)。放出された胞子は薄膜を脱ぎ捨て、べん毛の回転によって超高速で運動する遊走子となる。遊走子は栄養分が豊富な環境に辿り着くと運動を停止して、発芽し、再び菌糸状の生育を開始する。
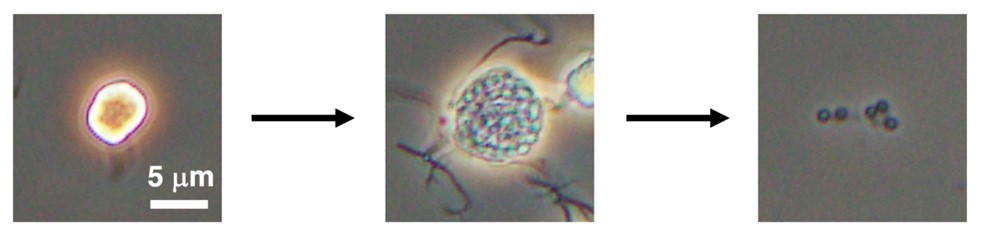
図2. 位相差顕微鏡で観察した胞子嚢開裂の過程
水に浸された直後、胞子嚢は光学顕微鏡下で光っているように見える(写真左)が、次第に胞子嚢膜が透明になり、内部が透けて見えるようになる(写真中央)。その後、胞子嚢膜が破れ胞子が放出される(写真右:放出された胞子の一部)。
研究成果
希少放線菌アクチノプラネス・ミズーリエンシスが形成する胞子嚢の開裂に着目し、その分子メカニズムの解明を目指して研究が行われました。まず、胞子嚢内部の胞子が遊走子よりも高い耐熱性を持つことを利用し、胞子嚢に水がかかっても開裂しない変異株(M-12株)が取得されました。さらにM-12株のゲノム解析等により、アンチシグマ因子(注3)SipAをコードする遺伝子に生じた変異が、この変異株の表現型の原因であることがわかりました。次に、M-12株をもとに、水がかかると正常に胞子嚢が開裂する抑圧変異株(注4)が取得され、シグマ因子(注3)SsdAをコードする遺伝子に生じた変異がこの表現型の原因であることを示されました。以上より、SipAとSsdAからなるシグマ/アンチシグマ因子システムの存在が明らかになり、これが胞子嚢の開裂を制御していることが示唆されました。
アンチシグマ因子SipAとシグマ因子SsdAのアミノ酸配列から、SipAはアンチシグマ因子ドメインに加えてアンチ・アンチシグマ因子(注5)に相同性があるドメインを持ち、SsdAもシグマ因子ドメインに加えてアンチ・アンチシグマ因子に相同性があるドメインを持っていることがわかりました。このようなドメイン構成をもつシグマ因子やアンチシグマ因子はこれまでに報告がなく、SipAとSsdAは新しい制御様式を有するシグマ/アンチシグマ因子システムを構成していると考えられました。アンチシグマ因子SipAとシグマ因子SsdAの結合様式が詳細に解析された結果、両者はSipAのアンチシグマ因子ドメインとSsdAのアンチ・アンチシグマ因子ドメイン間の結合を介して相互作用することがわかり、アンチ・アンチシグマ因子ドメインがシグマ因子とアンチシグマ因子間の相互作用を仲介することが明らかになりました。
また、胞子嚢が正常に開裂する別の抑圧変異株の解析より、二成分制御系(注6)の構成因子RsdKをコードする遺伝子に生じた変異がこの抑圧変異株の表現型の原因であることがわかりました。RNAシーケンス解析(注7)の結果、ヒスチジンキナーゼであるRsdKと応答制御因子RsdRからなる二成分制御系をコードする遺伝子の転写は、SipAとSsdAからなるシグマ/アンチシグマ因子システムの制御下にあることが明らかになりました。RsdKとRsdRの機能を調べたところ、この二成分制御系が胞子嚢の開裂を抑制し、水がかかっても胞子嚢が一斉に開裂しないようにコントロールしていることが明らかになりました。さらに、シグマ因子SsdAをコードする遺伝子を欠失させた変異株(ssdA破壊株)の解析により、ssdA破壊株の遊走子は野生株の遊走子と比べて酸化ストレスに対する耐性が低下していることがわかりました。以上の結果より、SipAとSsdAからなるシグマ/アンチシグマ因子システムは、胞子嚢開裂と胞子のストレス耐性に関わる遺伝子を包括的に制御していることが示唆されました。本研究により、希少放線菌アクチノプラネス・ミズーリエンシスでは、休眠状態と考えられてきた胞子において一部の遺伝子が発現し、休眠状態からの覚醒の抑制やストレス耐性を担っていることが示唆されました(図3)。
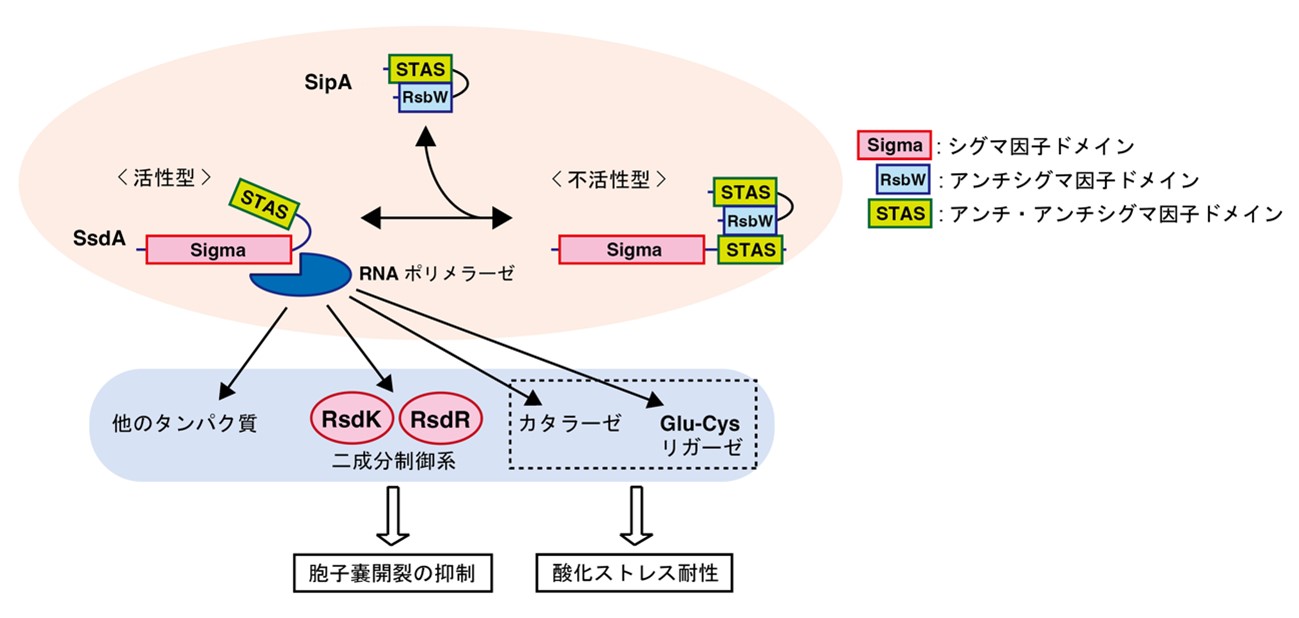
図3. 希少放線菌アクチノプラネス・ミズーリエンシスにおける胞子嚢開裂とストレス耐性の制御モデル
社会的意義
細菌における胞子の形成メカニズムはモデル微生物を用いて精力的に研究されてきました。一方、休眠状態にあると考えられる胞子が環境の変化を感知し、覚醒する分子メカニズムについては不明な点が多く残されています。本研究では、希少放線菌の胞子において、一部の遺伝子を発現させ休眠状態からの覚醒を抑制する制御システムの存在が示唆されました。この発見は、「胞子の生物学」において新しい概念を提供するものであり、病原性細菌の胞子の休眠や覚醒に関する研究にも一石を投じるものです。
発表雑誌
- 雑誌
- Nature Communications 14, Article number: 8483 (2023)
- 題名
- A unique sigma/anti-sigma system in the actinomycete Actinoplanes missouriensis
- 著者
- Takeaki Tezuka#,*, Kyota Mitsuyama#, Risa Date#, and Yasuo Ohnishi* (#共同筆頭著者, *共同責任著者)
- DOI
- https://doi.org/10.1038/s41467-023-44291-y
- URL
- https://www.nature.com/articles/s41467-023-44291-y
研究助成
本研究は、日本学術振興会科学研究費補助金(課題番号: JP26252020、JP18H02122、JP17K07711、JP20K05781、JP19H05685、JP16H06279)、A3 Foresight Programの支援を受けて行われました。
用語解説
- 注1 胞子嚢
内部に胞子を含む袋状の構造物。希少放線菌アクチノプラネス・ミズーリエンシスでは、1つの袋状の構造に100-200個程度の胞子を内包する胞子嚢が形成される。 - 注2 遊走子
べん毛により運動する胞子。希少放線菌アクチノプラネス・ミズーリエンシスでは、胞子嚢から放出された胞子はべん毛を持ち、水中を高速で運動するが、栄養増殖に適した環境では運動を停止し、発芽して菌糸生長を開始する。 - 注3 シグマ因子とアンチシグマ因子
シグマ因子はRNAポリメラーゼの構成因子であり、ゲノムDNA上で転写が開始される位置を決定するタンパク質。RNAポリメラーゼはゲノムDNAを鋳型として、リボヌクレオチドを重合させてRNAを合成する酵素複合体であり、RNAポリメラーゼと結合したシグマ因子は特異的なプロモーター配列を認識して結合することで、転写される遺伝子を決定する。アンチシグマ因子は、シグマ因子に結合することでRNAポリメラーゼ複合体とシグマ因子の結合を阻害するタンパク質。アンチシグマ因子が特定のシグマ因子に結合することで、シグマ因子によりプロモーター配列が認識される遺伝子の転写が抑制される。 - 注4 抑圧変異株
突然変異により生じた効果が、ゲノム上の別の位置に生じた突然変異により抑制された変異株。本研究では、sipA遺伝子に生じた突然変異によりM-12株が形成する胞子嚢の開裂が抑制されたが、さらにssdA遺伝子あるいはrsdK遺伝子に突然変異が生じることで胞子嚢が正常に開裂する抑圧変異株が得られた。 - 注5 アンチ・アンチシグマ因子
アンチシグマ因子に結合するタンパク質。アンチ・アンチシグマ因子が特定のアンチシグマ因子に結合することで、アンチシグマ因子はシグマ因子に結合することができなくなる。その結果、シグマ因子はRNAポリメラーゼと結合することが可能になり、特定の遺伝子の転写が開始される。 - 注6 二成分制御系
ヒスチジンキナーゼと応答制御因子の2つのタンパク質で構成される細胞応答の制御システム。ヒスチジンキナーゼは環境の変化を感知すると自己リン酸化し、このリン酸基がパートナーである応答制御因子に転移することで応答制御因子が活性化される。典型的な二成分制御系の応答制御因子は、リン酸化されることでゲノムDNAの特定の配列を認識して結合し、遺伝子の発現を活性化する。これにより、環境の変化に対する細胞の応答が誘導される。 - 注7 RNAシーケンス解析
次世代シーケンサーを用いて細胞から抽出したRNAを解析する手法。転写産物を網羅的、定量的に解析するトランスクリプトーム解析手法の1つ。次世代シーケンサーは数百万塩基対以上のDNA分子の配列を同時に決定できる解析装置であり、培養条件の異なる細胞を用いてRNAシーケンス解析を行うことで転写産物の量がどのように変化したかを網羅的に決定することができる。また、野生株と遺伝子破壊株の細胞を用いてRNAシーケンス解析を行うことで、両株において転写産物の量が異なる遺伝子を決定することができる。
問い合わせ先
<本研究に関するお問い合わせ>
東京大学 大学院農学生命科学研究科 応用生命工学専攻
醗酵学研究室 教授 大西 康夫(おおにし やすお)
Tel: 03-5841-5123 Fax: 03-5841-8021
E-mail: ayasuo[アット]g.ecc.u-tokyo.ac.jp
醗酵学研究室HP: https://www.hakko.bt.a.u-tokyo.ac.jp
東京大学大学院農学生命科学研究科・農学部
事務部 総務課総務チーム 総務・広報情報担当(広報情報担当)
電話:03-5841-8179, 5484、Eメール:koho.a[アット]gs.mail.u-tokyo.ac.jp
FAX:03-5841-5028
※[アット]を@に変えてください。