細菌べん毛の回転を停止させる新規タンパク質の発見
発表のポイント
- 希少放線菌の遊走子のべん毛回転を停止させる新規タンパク質FtgAを発見しました。
- FtgAの作用点を明らかにするとともに、遊走子が栄養源を感知して運動を停止する分子機構を提唱しました。
- 今回の発見は、高性能ナノマシンである細菌べん毛モーターの分子機構解明の新たな糸口になると期待されます。
発表概要
東京大学大学院農学生命科学研究科・醗酵学研究室の大西康夫教授・手塚武揚助教(現・微生物エコテクノロジー社会連携講座特任講師)の研究グループは、希少放線菌における形態分化の分子機構について長年研究してきました(参考URL 1-3)。希少放線菌Actinoplanes missouriensis(アクチノプラネス・ミズーリエンシス)は100-200個程度の胞子を内包した胞子嚢(注1)を形成します。胞子嚢に水がかかると胞子が覚醒し、胞子嚢から放出されます。放出された胞子はべん毛を回転させることで水中を高速で運動する遊走子(注2)となります。遊走子は走化性(注3)を有しており、自身の生育に適した栄養豊富な環境に到達すると運動を停止し、発芽します。今回、遊走子の走化性を制御していると予想される遺伝子群の研究から、遊走子の運動の停止に関与する遺伝子が見出されました。この遺伝子の破壊株では、栄養源を感知しても遊走子が運動を停止せず、発芽した後も細胞が運動し続けるようになりました。この遺伝子がコードするタンパク質は、べん毛モーターの構成タンパク質の1つに特異的に結合しますが、このべん毛モータータンパク質にアミノ酸置換が生じた変異株では、遊走子が栄養源を感知しても運動を停止しませんでした。一方、運動停止に関わるタンパク質は、べん毛モーターの構成タンパク質に加えて、化学物質を感知すると予想される走化性受容体を構成する2つのタンパク質にも結合することが明らかになりました。以上の結果から、今回、希少放線菌で発見されたタンパク質は、遊走子の運動中は走化性受容体の構成因子としてタンパク質複合体に係留されている一方、栄養源となる化学物質が走化性受容体に結合するとこの複合体から解離し、べん毛モーターの構成タンパク質に結合してべん毛の回転を停止させる、という作用モデルを提唱しました。細菌のべん毛モーターは非常に高性能なナノマシンとして知られており、本研究で発見されたべん毛回転のブレーキとして働くタンパク質は、べん毛モーターの分子機構解明の新たな糸口になることが期待されます。
参考URL
- 注目を集める希少放線菌の不思議に迫る研究(東京大学大学院農学生命科学研究科 研究成果 2017/06/08)
- 注目を集める希少放線菌の不思議に迫る研究:遊走子線毛の発見と機能解明(東京大学大学院農学生命科学研究科 研究成果 2019/07/10)
- 細菌胞子の休眠と覚醒の制御に関する新たな知見:放線菌の胞子嚢胞子は完全には休眠していない?(東京大学大学院農学生命科学研究科 研究成果 2023/12/26)
発表内容
背景
一部の細菌は、べん毛の回転による運動性を示します。運動性は、細胞が貧栄養な環境から栄養豊富な環境に移動することを可能にするなど、細菌における重要な生存戦略の1つです。また、細菌のべん毛モーターは、エネルギー変換効率がほぼ100%の非常に高性能なナノマシンであるため、その回転運動のメカニズムを分子レベルで解明することは、超高効率微小ナノマシンの開発につながると期待され、これまで多くの研究がなされてきた重要な研究領域です。
希少放線菌Actinoplanes missouriensis(アクチノプラネス・ミズーリエンシス)は、菌糸細胞の伸長・分岐により増殖しますが、貧栄養な環境では100-200個程度の胞子を内包した胞子嚢(注1)を形成して休眠状態になります。胞子嚢は水がかかると表層の膜(胞子嚢膜)が破れ、内部の胞子を放出します。胞子嚢から外部に放出された胞子は遊走子(注2)となり、1時間程度、水中を高速で移動することができます。一方で、遊走子は走化性(注3)を有しており、増殖に適した環境に到達すると運動を停止し、発芽して菌糸細胞の伸長を開始します(図1)。このように、希少放線菌A. missouriensisは、細菌としては極めて高度な形態分化能(異なった形態・能力を有する細胞を生じる能力)を有しており、A. missouriensisの生活環において遊走子は、ごく短時間のみ運動性を獲得する特異な状態の細胞と位置づけられます(図2左)。
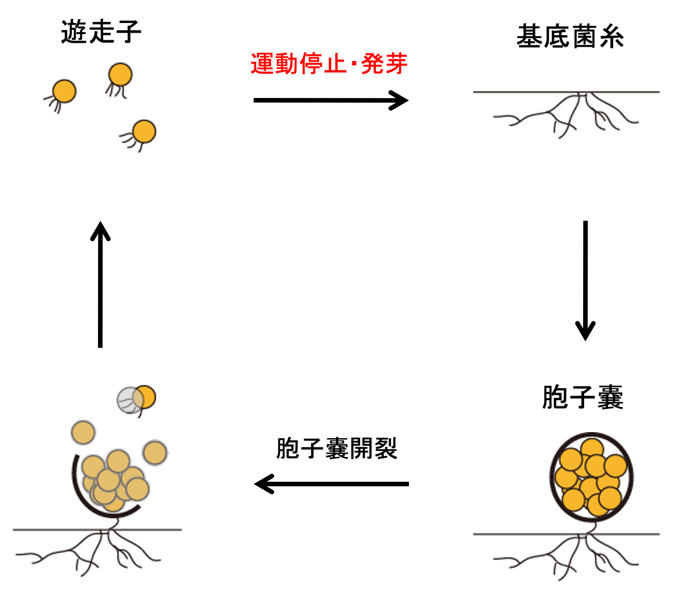
図1:希少放線菌A. missouriensisの生活環
栄養分が枯渇すると、栄養増殖細胞である基底菌糸から、100-200個程度の胞子を内包した胞子嚢が形成される。水に浸されると胞子嚢の膜が破れ、胞子が水中に放出される。胞子は薄膜を脱ぎ捨て、べん毛の回転によって高速で運動する遊走子となる。遊走子は栄養分が豊富な環境にたどり着くと運動を停止して発芽し、再び菌糸状の生育を開始する。
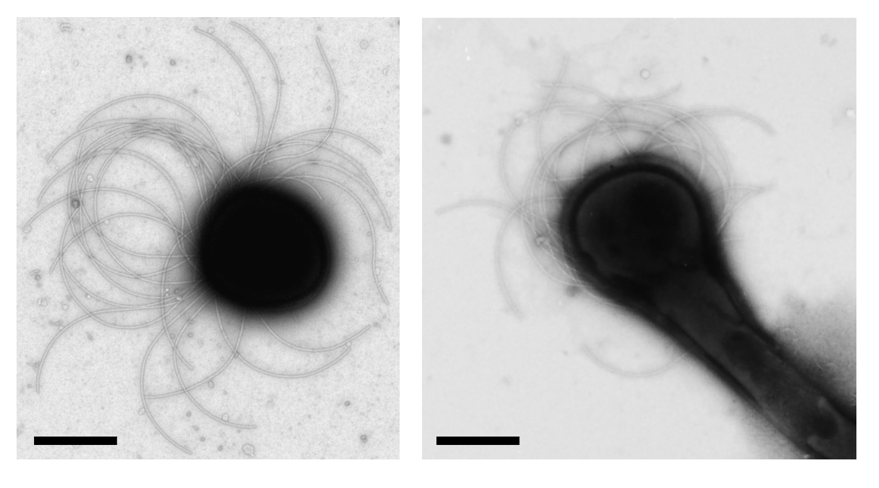
図2:透過型電子顕微鏡で観察した遊走子と発芽細胞
(写真左)遊走子をネガティブ染色した観察像。(写真右)遊走子を栄養豊富な液体培地に懸濁し、30℃で8時間培養した後、遊走子と同様に観察した。スケールバーはいずれも1マイクロメートル。
研究成果
本研究では、希少放線菌A. missouriensisが形成する遊走子が、栄養源を感知して運動を停止することに着目し、その分子メカニズムの解明を目指しました。まず、栄養豊富な液体培地に遊走子を懸濁して発芽させ、透過型電子顕微鏡で観察したところ、発芽した細胞に遊走子と同じくべん毛が観察されたことから、遊走子は運動を停止する際にべん毛を脱落させるのではなく、何らかの機構で回転を停止させていることがわかりました(図2右)。一方、走化性の制御に関与すると予想される遺伝子群 (che cluster-1; 以下che-1) の機能解析において、che-1の欠損株では、遊走子が発芽した後もべん毛が回転し続けることを発見しました。che-1には10個の遺伝子が含まれるため、遺伝子破壊による解析をさらに進めたところ、機能未知タンパク質をコードする遺伝子ftgAを破壊すると、遊走子が発芽した後もべん毛回転が停止しなかったことから、この機能未知タンパク質がべん毛回転の「ブレーキタンパク質」として機能することが示唆されました。
FtgAタンパク質の機能をさらに解析するため、ftgA遺伝子の過剰発現株を作製したところ、この過剰発現株の胞子嚢から放出された胞子には全く運動性がないことがわかりました。当初はブレーキタンパク質がたくさん作られるために、べん毛回転が常に止められていると考えていましたが、後になってこの胞子にはべん毛が形成されていないことがわかりました。次に、この過剰発現株をもとに、胞子嚢から放出された胞子が正常に水中を運動する抑圧変異株(注4)を取得しました。さらに解析を進めたところ、この変異株においては、べん毛モーターの構成因子であるFliNをコードする遺伝子に生じた変異により、正常に胞子べん毛が形成され、運動能が回復したことが明らかになりました。また、fliN遺伝子に生じたこの変異を野生株に導入したところ、ftgA破壊株と同様、遊走子が発芽した後もべん毛回転が停止しないことがわかりました。これらの結果から、FtgAはべん毛モーターに直接作用することで、べん毛回転を停止させていることが示唆されました。(FliNはべん毛形成にも関与しているため、FtgAの過剰生産によって、べん毛が形成されなくなることも説明できます。)
そこで、FtgA-FliN間の相互作用を解析したところ、FtgAはFliNタンパク質に結合することがわかりました。さらに、当初の予想に反して、FtgAはアミノ酸配列に置換が生じた変異型FliNタンパク質にも野生型FliNと同様に結合することがわかりました。このことから、(i) FtgAはべん毛モーターを構成するFliNに結合することでべん毛回転を停止させること、(ii) 変異型FliNを構成因子とするべん毛は、FtgAがFliNに結合してもべん毛回転が停止しないこと、が明らかになりました。通常では、FtgAの結合によって引き起こされるFliNの構造変化がべん毛モーターの回転を停止させるが、変異型FliNでは、この構造変化が正常に起こらないために、べん毛モーターの回転を停止できないものと推測されます。
FtgAはche-1中の遺伝子にコードされていることから、che-1の他の遺伝子がコードするタンパク質とFtgAとの相互作用を解析したところ、FtgAはCheA1およびCheW1-2の2つのタンパク質と相互作用することが明らかになりました。CheA1とCheW1-2はいずれも、特定の化学物質の濃度勾配を感知する走化性受容体の構成因子であることから、FtgAもこのタンパク質複合体の構成因子である可能性が示唆されました。以上の結果より、遊走子における運動停止の分子機構として、(1) 遊走子が水中を運動している際には、FtgAは走化性受容体の構成因子としてCheA1およびCheW1-2に結合している、(2) 走化性受容体に何らかの化学物質が結合すると、FtgAは複合体から解離してべん毛モーターを構成するFliNに結合し、べん毛回転を停止させる、という制御モデルが提唱されました(図3)。
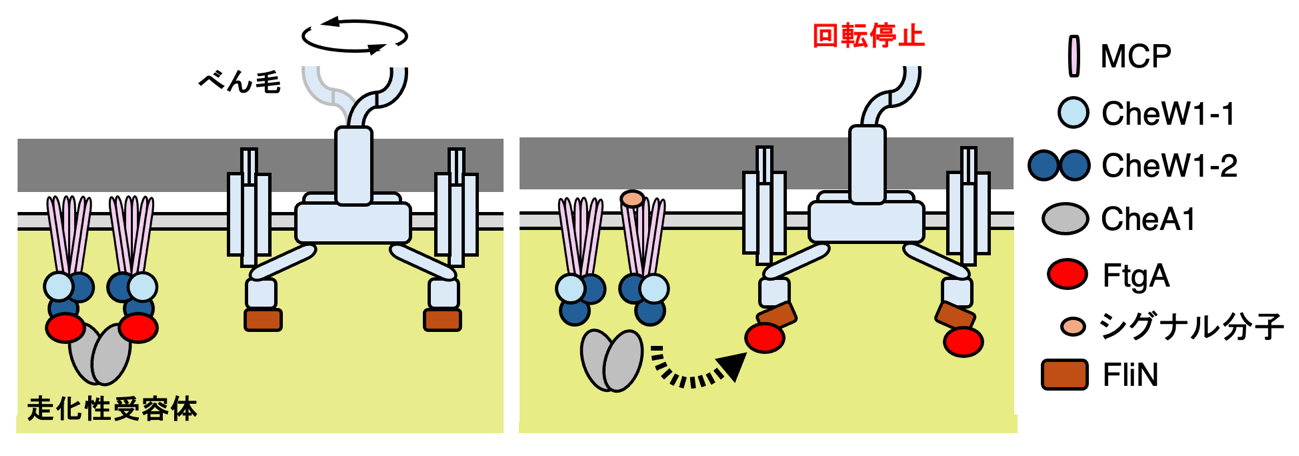
図3:希少放線菌A. missouriensisにおける遊走子の運動停止の制御モデル
社会的意義
細菌のべん毛モーターはエネルギー変換効率がほぼ100%の高性能ナノマシンであり、その分子機構の解明は超高効率微小モーターの開発につながると期待され、これまで多くの研究がなされてきました。大腸菌や枯草菌を含むいくつかの運動性細菌において、べん毛回転を抑制するタンパク質が複数発見されていますが、今回発見された「ブレーキタンパク質」は、その起源や作用機構がこれら既知のタンパク質とは異なると考えられます。遊走子べん毛の回転を停止させる新規な「ブレーキタンパク質」は、その作用機構の解明を通して、べん毛モーターの分子機構解明の新たな糸口になることが期待されます。
発表者
東京大学大学院農学生命科学研究科 応用生命工学専攻
加藤 大夢 修士課程(当時)
種村 裕幸 修士課程(当時)
木村 知宏 博士課程(当時)
勝山 陽平 准教授
手塚 武揚 特任講師
大西 康夫 教授
論文情報
- 雑誌
- Communications Biology 7, Article number: 1405 (2024)
- 題名
- Molecular mechanism of flagellar motor rotation arrest in bacterial zoospores of Actinoplanes missouriensis before germination
- 著者
- Hiromu Kato, Hiroki Tanemura, Tomohiro Kimura, Yohei Katsuyama, Takeaki Tezuka *, and Yasuo Ohnishi*(*共同責任著者)
- DOI
- 10.1038/s42003-024-07104-6
- URL
- https://www.nature.com/articles/s42003-024-07104-6
研究助成
本研究は日本学術振興会科学研究費補助金(課題番号: JP26252010、JP18H02122、JP17K07711、JP20K05781、JP19H05685)、A3 Foresight Programの支援を受けて行われました。
用語解説
- 注1 胞子嚢
内部に胞子を含む袋状の構造物。希少放線菌A. missouriensisでは、1つの袋状の構造に100-200個程度の胞子を内包する胞子嚢が形成される。 - 注2 遊走子
べん毛により運動する胞子。希少放線菌A. missouriensisでは、胞子嚢から放出された胞子がべん毛を持ち、水中を高速で運動するが、栄養増殖に適した環境では運動を停止し、発芽して菌糸生長を開始する。 - 注3 走化性
細胞が自身の周囲に存在する特定の化学物質の濃度勾配を感知し、それに対して誘因あるいは忌避行動を示す現象。 - 注4 抑圧変異株(サプレッサー変異株)
遺伝子の変異により生じた効果が、ゲノム上の別の位置に生じた突然変異により抑制された変異株。本研究では、ftgA遺伝子の過剰発現により胞子のべん毛合成が抑制されたが、fliN遺伝子に突然変異が生じることで胞子のべん毛が正常に合成される抑圧変異株が得られた。
問い合わせ先
東京大学大学院農学生命科学研究科 応用生命工学専攻 醗酵学研究室
教授 大西 康夫(おおにし やすお)
E-mail:ayasuo[at]g.ecc.u-tokyo.ac.jp
醗酵学研究室HP:https://www.hakko.bt.a.u-tokyo.ac.jp
東京大学大学院農学生命科学研究科 応用生命工学専攻
微生物エコテクノロジー社会連携講座
特任講師 手塚 武揚(てづか たけあき)
E-mail: atezuka[アット]g.ecc.u-tokyo.ac.jp
※上記の[at]は@に置き換えてください。